Voltage-Gated Proton Channel HVCN1 in B-Cell Activation
Study on HVCN1 co-localization with BCR and its role in B-cell activation, phosphorylation, and proton conductance modulation in vivo. Investigation of HVCN1 deficient mouse phenotype and impact on BCR signaling strength.

Voltage-Gated Proton Channel HVCN1 in B-Cell Activation
E N D
Presentation Transcript
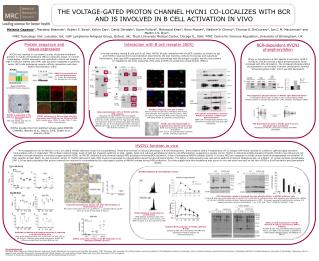
Time: 0 Time:60’ Time:60’ Time:60’ THE VOLTAGE-GATED PROTON CHANNEL HVCN1 CO-LOCALIZES WITH BCR AND IS INVOLVED IN B CELL ACTIVATION IN VIVO Melania Capasso1, Mandeep Bhamrah1, Robert S. Boyd1, Kelvin Cain1, David Dinsdale1, Karen Pulford2, Mahmood Khan4, Boris Musset3, Vladimir V. Cherny3, Thomas E. DeCoursey3, Ian C.M. MacLennan4 and Martin J.S. Dyer1. 1MRC Toxicology Unit, Leicester, UK; 2LRF Lymphoma Antigens Group, Oxford, UK; 3Rush University Medical Center, Chicago IL, USA; 4MRC Centre for Immune Regulation, University of Birmingham, UK Protein sequence and tissue expression Interaction with B cell receptor (BCR) BCR-dependent HVCN1 phosphorylation In human primary resting B cells and B cell lines, HVCN1 directly interacted with the BCR complex, as shown by Igb and HVCN1 reciprocal immunoprecipitation experiments. We also found by confocal microscopy and subcellular fractionation, that upon BCR engagement the channel was internalised with the antigen receptor and the two proteins co-migrated to the endo-lysosomal, MHC class II (MHC-II) containing compartments (MIICs). HVCN1 was isolated in a proteomic survey of plasma membrane proteins in mantle cell lymphoma (MCL) in leukemic phase. In normal lymphocytes, HVCN1 expression was restricted to the B-cell lineage, high in follicular mantle zone (FM) cells but down-regulated in germinal center (GC) cells undergoing receptor affinity maturation and class-switch recombination (CSR). When re-introduced in a HEL-specific B cell clone, LK35.2 HyHEL10, HVCN1 showed a basal phosphorylation which increased with HEL stimulation. The increased phosphorylation corresponded to an increase in proton conductance, termed “enhanced gating mode” and it was PKC dependent. HVCN1 INTRACELLULAR a b GC HVCN1 is expressed in B cells and granulocytes T= T cells, M= monocytes, G= granulocyte, B= B cells EXTRACELLULAR control FM Sosui prediction for HVCN1 human sequence. PKC-dependent phosphorylation of HVCN1 upon BCR stimulation 32P phosphorylation assay of LK35.2 cells overexpressing HVCN1. Phosphorylation is strongly abrogated by a PKC inhibitor, GF109203X (GFX). c BCR stimulated HVCN1co-migrates with IgM to MIICs Human primary B cells were stimulated with F(ab’)2 FITC-anti-IgM. The lysosome marker Lamp1, MHC-II molecule HLA-DR and MHC-II like molecule HLA-DM indicate MHC-II containing compartments. Immunoprecipitation (IP)of HVCN1 with the BCR complex Reciprocal IP experiments with myc-tagged, overexpressed HVCN1 and BCR component Igb (a,b) in A20 D1.3 cells, specific for hen egg lysozyme (HEL). The interaction does not change upon BCR stimulation with HEL (b). Co-IP of endogenous HVCN1 with Igb in GRANTA 519 cells. In this cell line HVCN1 is present as two isoforms (arrows, c). Sub-cellular fractionation confirms HVCN1 localisation to MIICs upon BCR stimulation BJAB cells overexpressing HVCN1 were analysed at steady state and upon IgM stimulation. HVCN1 is expressed in FM cells but downregulated in GC cells IHC with HVCN1 anti-sera of tonsil sections. HVCN1 expression in MZ overlaps with IgD expression and CD20 but not CD3 and CD68 IgD= mantle zone; CD20= B cells; CD3= T cells; CD68= macrophages PKC-dependent increase in HVCN1 proton current amplitude upon stimulation Properties of HVCN1 conductance change with PMA stimulation HVCN1 and internalised IgM co-localisation by electron microscopy BJAB overexpressing HVCN1 upon IgM stimulation. 5nm and 10 nm gold particles represent HVCN1 and IgM, respectively. HVCN1 encodes the first identified voltage-gated PROTON CHANNEL (Ramsey et al., Nature, 2006; Sasaki et al., Science, 2006) HVCN1 function in vivo To investigate the role of HVCN1 in vivo, we used a HVCN1-deficient mouse line (unpublished). These mice showed no obvious phenotype in B-cell development. Immunization with a T-independent (TI-2) antigen (NP-Ficoll) resulted in a defect in IgM and IgG3 secretion. Immunization with a T-dependent (TD) antigen induced similar levels of IgM but impaired switching to IgG1, IgG2b, IgG3, and reduced extrafollicular plasma cell differentiation, suggesting a pivotal role for HVCN1 in determining BCR-signaling strenght. HVCN1 may influence B-cell activation through alteration of reactive oxygen species (ROS), as HVCN1-deficient B cells showed reduced ROS production following BCR activation, a sign of suboptimal NADPH oxidase activity. In B cells, ROS are necessary to inhibit protein tyrosine phosphatases via oxidation of their catalitic domain (Reth, M., Nat Immunol, 2002). In HVCN1-deficient B cells, ROS levels corresponded to reduced BCR-induced tyrosine phosphorylation. The defect in BCR-signaling was rescued by addition of Sodium Stibogluconate, an inhibitor of protein tyrosine phosphatase SHP-1. It has been postulated that proton channels are required to counterbalance the electrogenic activity of NADPH oxidase during ROS production. Our data suggest that this mechanism also occurs in vivo and shed new light on the role of ROS in B cell activation and downstream effects. TI-2: NP-Ficoll HVCN1 deficient B cell defects in vitro: TD: NP-KLH BCR stimulation results in impaired tyrosine phosphorilation in HVCN1 deficient cells Purified B cells were stimulated with 20 mg/ml F(ab’)2 anti-IgM for indicated times. BCR-signaling was inhibited by addition of 15 mM of a NADPH oxidase inhibitor, diphenyleneiodonium (DPI). The graph represents densitometric analysis of 4 separate phospho-tyrosine immunoblots. * p= 0.031 and **p= 0.00488 by Student’s t test. Impaired extrafollicular plasma cell (PC) differentiation in HVCN1 deficient mice Mice were immunized with 100 mg NP-KLH s.c., boosted with 25 mg NP-KLH i.p. at day 8 and culled at day 12. fold increase HVCN1 deficient B cells defect in proliferation CFSE-labeled, purified B cells were stimulated with 1.5 mg/ml F(ab’)2 anti-IgM/ 20 ng/ml IL-4 and cultured for 48h. Defect in BCR stimulation in HVCN1 deficient B cells is rescued by a SHP-1 inhibitor Purified B cells were stimulated with 20 mg/ml F(ab’)2 anti-IgM in the presence or absence of 5 mM Sodium Stibogluconate, an inhibitor of SHP-12. Reduced ROS production in HVCN1 deficient B cells Isolated B cells were stimulated with PMA and analysed for ROS production by chemiluminescence. Antibody production and isotype switching is impaired in HVCN1-deficient mice Mice were immunized with 100 mg NP-Ficoll i.p. and 100 mg NP-KLH s.c. Antibody titers were analysed by ELISA. All differences significant by Mann-Whitney test. Affinity maturation of IgG1 anti-NP antibodies is not affected by HVCN1 absence Acknowledgments We thank Paolo Salomoni, Pierluigi Nicotera and Gerry Cohen laboratories for reagents and scientific discussions (MRC Toxicology Unit, Leicester, UK); Elena Vigorito and Susan Kohlhaas (The Babraham Institute, Cambridge, UK) for useful discussions, Kenneth Smith laboratory (Cambridge Institute for Medical Research, University of Cambridge, Cambridge, UK) for reagents and scientific discussions, Facundo Batista (CRUK, London, UK) and Andrew Knight (Newcastle University, Newcastle, UK) for reagents. This work is funded by Lymphoma Research Found and the Medical Research Council.