Download

1 / 1
10 likes | 106 Views
Explore the journey from Triosephosphate Isomerase to Kealases as we engineer A-TIM with wider substrate range. Utilizing Oulu campus tools, we design new enzymes by mutating regions for improved catalysis.

E N D
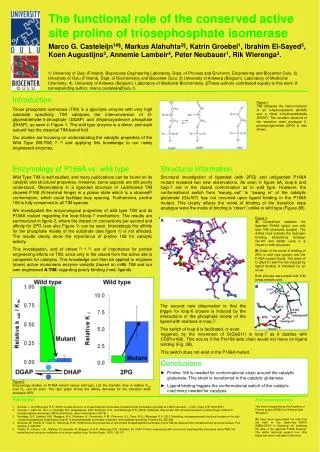
Figure 3b Figure 3a Figure 2 Figure 1 Input and out put of the project. In blue are the mutagenesis experiments, in yellow the analytical tools available on the Oulu campus, and in purple the final goal. Results so far: A-TIM, Binders, Novel compounds. A Novel Sulphonate binds A-TIM (1.7 Å). The red regions in A-TIM are targeted for site-directed mutagenesis. In grey is shown the backbone and in white the catalytic residues. Both pictures were made with ICM (www.molsoft.com). Connolly surface picture of the new extended groove (at 1.06Å). The side chains of Lys13 (catalytic residue) and Lys239 (rim residue) are shown in transparent mode. Regions of negative electrostatic potential are colored red, and positive electrostatic potential are colored blue. Wild type TIM (Wt-TIM) has high substrate specificity, and only catalyses the interconversion of the α-hydroxykentone DHAP and the α-hydroxyaldehyde D-GAP. The transition state analogue, 2-phosphoglycolate (2-PG), can be seen on the right. Red box: reactive head group (R)Green box: anchor moiety (A) Creating new enzymes - From Triosephosphate Isomerase to Kealases Marco G. Casteleijn1, Markus Alahuhta2, Mikko Salin2, Matti Vaismaa3, Ritva Juvani3, Nanna Alho3, Sampo Mattila3,Marja Lajunen3, Jouni Pursiainen3, Rik K. Wierenga2#, Peter Neubauer1#. 1) University of Oulu (Finland), Bioprocess Engineering Laboratory, Dept. of Process and Environm. Engineering; 2) University of Oulu (Finland), Dept. of Biochemistry; 3) University of Oulu (Finland), Dept. of Chemistry. # corresponding author:rik.wierenga@oulu.fi, peter.neubauer@oulu.fi Introduction R Triose phosphate isomerase (TIM) is a glycolytic enzyme with very high substrate specificity (fig 1). The wild type enzyme is a dimer, and each subunit has the classical TIM-barrel fold[1]. Several loop modifications resulted in a monomeric enzyme (ml1TIM)[2,3] which remains active, although less so than the wild-type. Subsequently this has resulted in in a new variant (A-TIM) with a much more extensive binding[4,5]. Our current studies attempt to change the substrate specificity of monomeric TIM. A The right starting point The goal: Kealases Our aim is to build a platform of TIM variants (Kealases) with a widened substrate range. Therefore we established by rational design a library of TIM variants, which resulted in a candidate for further development: A-TIM (fig. 2). A break through observation is that newly designed ligands bind into this pocket (fig. 3b; example). The use of the analytical tools available on the Oulu campus have shown to drive protein engineering development and the creation of tailored ligands for the new active site of A-TIM. Overall, A-TIM is an ideal test case for directed evolution approaches aiming at fine tuning its catalytic properties, and a very suitable starting protein for a bio-catalytic platform for the following reasons: Wild type and binding studies are focused on the binding and catalysis dynamics of the active site. Loop 7 is responsible for initiating conformational changes within the active site by “capturing” (together with loop 8) the anchor moiety (A; fig. 1) of the ligand[7]. Active site New binding groove • Small size, monomeric protein (very suitable for NMR; easily crystallized) • Highly expressed in E. coli as a soluble protein • Stable molecule, which tolerates many mutations • The current set of X-ray structures suggest that it is a “flexible” molecule, and therefore an ideal starting point for directed evolution experiments • TIM does not require any cofactors • In the closed conformation the binding site is an extended groove • Its wild type precursor is the extensively studied TIM[6] In figure 3 the understanding of the structure and its catalytic machinery is summarized in respect to the creation of new enzymes which interconvert α-hydroxykentones and α-hydroxyaldehydes. Shown are the new binding groove and the binding of a novel ligand for TIM. Structures of A-TIM liganded with 2-PG (fig.1) and a suicide inhibitor Bromo-hydroxyacetone phopsphate (BHAP), and NMR studies with original substrates (fig. 1 and 4) show that A-TIM still has a competent active site. A-TIM variants, derived from randomized mutagenesis, that can convert new ligands will be called Kealases. DHAP D-GAP Figure 4 1-D NMR spectrum of the conversion of D-GAP into DHAP under the influence of ATIM Conclusions • Monomeric TIM is a good scaffold for enzyme engineering. • The newly engineered A-TIM has a competent active site and binds novel ligands. References Acknowledgements This work is supported by the Academy of Finland (project 53923 & 117874). The authors would also like to thank Prof. Dr. Koen Augustyns from the University of Antwerp (Belgium) for providing us with Bromohydrogen-acteone phosphate (BHAP), and Ville Ratas2 for his technical support. • Kursula, I., and Wierenga, R.K. (2003) Crystal structure of triosephosphate isomerase complexed with 2-phospho-glycolate at 0.83Å resolution. J. Biol. Chem 278, 9544-9551 • Thanki, N., Zeelen, J.Ph., Mathieu, M, Jaenicke, R, Abagyan, R.A.A., Wierenga, R.K., Schliebs, W. (1997) Protein engineering with monomeric triophosphate isomerase (monoTIM): the modeling and structure verification of a seven-residue loop. Protein Engin. 10(2), 159-167 • Thanki N, Zeelen JP, Mathieu M, Jaenicke R, Abagyan RA, Wierenga RK, Schliebs W Protein engineering with monomeric triosephosphate isomerase (monoTIM): the modelling and structure verification of a seven-residue loop Protein Eng. v10, p.159-167 • Norledge, B.V., Lambeir, A.M., Abagyan, R.A., Rottmann, A., Fernandez, A. M., Filimonov, V.V., Peter, M.G., Wierenga, R.K. 2001. Modelling, mutagenesis and structural studies on the fully conserved phosphate binding loop (loop-8) of triosephosphate isomerase: towards a new substrate specificity. Proteins 42, 383-389. • Markus Alahuhta, Mikko Salin, Marco G. Casteleijn, Kristian Kemmer, Peter Neubauer, Rik K. Wierenga. Structure-based enzyme engineering efforts with an inactive monomeric TIM variant: the importance of a single point mutation for generating a competent active site. Manuscript • http://www.oulu.fi/bioprocess/biocatalysis [Kealases and Biocat projects] • Casteleijn MG, Alahuhta M, Groebel K, El-Sayed I, Augustyns K, Lambeir AM, Neubauer P, Wierenga RK. Functional Role of the Conserved Active Site Proline of Triosephosphate Isomerase. Biochemistry. 2006 Dec 26;45(51):15483-15494