
Chromosomes: Structure and Function
E N D
Presentation Transcript
Chromosomes: Structure and Function MOLECULAR GENETICS (856)
Changes in Chromatin Structure • Polytene chromosomes are giant chromosomes found in certain tissues of Drosophila and some other organisms. • These large, unusual chromosomes arise when repeated rounds of DNA replication take place without accompanying cell divisions, producing thousands of copies of DNA that lie side by side. • Chromosomal puffs—localized swellings of the chromosome. Eachpuff is a region of the chromatin that has relaxed its structure, assuming a more open state. If radioactively labeled uridine is added to a Drosophila larva, radioactivity accumulates in chromosomal puffs, indicating that they are regions of active transcription. • Appearance of puffs at particular locations on the chromosome can be stimulated by exposure to hormones and other compounds that are known to induce the transcription of genes at those locations. This correlation between the occurrence of transcription and the relaxation of chromatin at a puff site indicates that chromatin structure undergoes dynamic change associated with gene activity.
Chromosomal puffs in condensed Drosophila chromosome show states of de-condensing in expressed regions
A several experiments indicating that chromatin structure changes with gene activity is sensitivity to DNase I, an enzyme that digests DNA. • The ability of this enzyme to digest DNA depends on chromatin structure: when DNA is tightly bound to histone proteins, it is less sensitive to DNase I, whereas unbound DNA is more sensitive to digestion by DNase I. • The results of experiments that examine the effect of DNase I on specific genes show that DNase sensitivity is correlated with gene activity. • For example, globin genes code for hemoglobin in the erythroblasts (precursors of red blood cells) of chickens. The forms of hemoglobin produced in chick embryos and chickens are different and are encoded by different genes (Fig.a). • No hemoglobin is synthesized in chick embryos in the first 24 hours after fertilization. If DNase I is applied to chromatin from chick erythroblasts in this first 24-hour period, all the globin genes are insensitive to digestion (Fig.b).
Changes in Chromatin Structure • From day 2 to day 6 after fertilization, after hemoglobin synthesis has begun, the globin genes become sensitive to DNase I, and the genes that code for embryonic hemoglobin are the most sensitive (Fig c). • After 14 days of development, embryonic hemoglobin is replaced by the adult forms of hemoglobin. The most sensitive regions now lie near the genes that produce the adult hemoglobins (Fig.d). • DNA from brain cells, which produce no hemoglobin, remains insensitive to DNase digestion throughout development (Fig.e).
Method: Sensitivity to DNase I was tested on different tissues and at different times in development DNase I sensitivity is correlated with the transcription of globin genes in erythroblasts of chick embryos. The U gene codes for embryonic hemoglobin; the D and A genes code for adult hemoglobin.
Changes in Chromatin Structure • In summary, when genes become transcriptionally active, they also become sensitive to DNase I, indicating that the chromatin structure is more exposed during transcription. What is the nature of the change in chromatin structure that produces chromosome puffs and DNase I sensitivity? • In both cases, the chromatin relaxes; histones loosen their grip on the DNA. One process that appears to be implicated in changing chromatin structure is acetylation, a reaction that adds chemical groups called acetyls to the histone proteins. • Enzymes called acetyltransferases attach acetyl groups to lysine amino acids at one end (called a tail) of the histone protein. • This modification reduces the positive charges that normally exist on lysine and destabilizes the nucleosome structure, and so the histones hold the DNA less tightly. Proteins taking part in transcription can then bind more easily to the DNA and carry out transcription.
Histones are subject to modification which affect gene regulation • Exposed regions of the histone are subject to modification • These modification influence the structure of chromatin and it regulator properties • Some modification • Acetylation • Methylation • Phosphorylation
How are chromosome organized in a nucleus Staining pattern results in different color specific for each chromosome Chromosomes occupy discrete territories in the cell nucleus Chicken nuclei
Features of human chromosome territories specific location of p and q arms and specific territories Transparent view Three dimensional view of chromsome territories Same specific territories for the active and inactive X chromosome
Where are gene-rich and gene-poor regions of chromosome located Gene-poor chromosome located at nuclear periphery So generally silent regions of chromosomes are located at the nuclear periphery Gene-rich chromosome located in nuclear interior
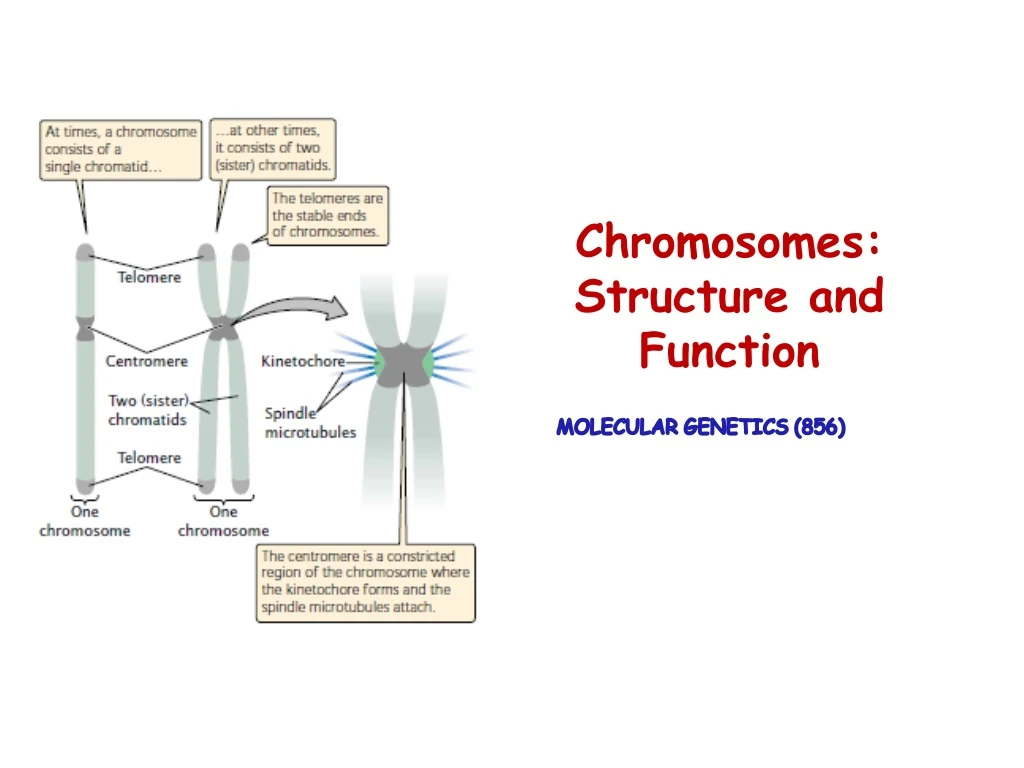
Essential Functional elements of Chromosomes • A functional chromosome has three essential elements: • A centromere • Apair of telomeres • Origins of replication
Chromosomes as functioning organelles Centromere Chromosome fragments that lack a centromere are lost in mitosis. • A constricted region of the chromosome where spindle fibers attach and is essential for proper movement of the chromosome in mitosis and meiosis . • The first centromeres to be isolated from Yeast (small, linear chromosomes).
Chromosomes as functioning organelles Centromere • Centromeric sequences are the binding sites for proteins that function as the kinetochore, a complex that assembles on the centromere and to which the spindle fibers attach. • Kinetochores play a central role in this process, by controlling assembly and disassembly of the attached microtubules and, through the presence of motor molecules, by ultimately driving chromosome movement.
Chromosomes as functioning organelles Centromere • In simple eukaryotes, the sequences that specify centromere function are very short. • Yeast Saccharomyces cerevisiae the centromere element (CEN) is about 110 bp long, comprising two highly conserved flanking elements of 9 bp and 11 bp and a central AT-rich segment of about 80–90 bp. • The centromeres of such cells are interchangeable - a CEN fragment derived from one yeast chromosome can replace the centromere of another with no apparent consequence. • In mammals, centromeres comprise hundreds of kilobases of repetitive DNA, some nonspecific and some chromosome-specific. Centromeres consist of particular sequences repeated many times. This nucleotide sequence is found in the point centromere of Saccharomyces cerevisiae.
Chromosomes as functioning organelles Centromere Variations in centromeric sequences • Diffuse Centromeres, spindle fibers attach along the entire length of the chromosome. • Localized centromeres; spindle fibers attach at a specific place on the chromosome. Appear constricted, but there also can be secondary constrictions at places that do not have centromeric functions. Classes of localized centromeres • Point centromeres • Smaller and more compact. DNA sequences are both necessary and sufficient to specify centromere identity and function in organisms with point centromeres) • Budding yeast (Saccharomyces cerevisiae) encompasses 125 bp of DNA. • Regional centromeres (large amounts of DNA and are often packaged into heterochromatin). • Most of the centromere is made up of short sequences of DNA that are repeated thousands of times in tandem. Within these repeats are “islands” of more complex sequence, primarily transposable element sequences.
Chromosomes as functioning organelles Centromere • Centromeric DNA shows remarkable sequences heterogeneity. • Universally marked by the presence of a centromere-specific variant of histone H3, generically known as CenH3 (the human form of CenH3 is named CENP-A). • At centromeres, CenH3/CENP-A replaces the normal histone H3 and is essential for attachment to spindle microtubules. • Depending on centromere organization different numbers of spindle microtubules can be attached.
Chromosomes as functioning organelles Centromere Human Centromeres: • 171 bp monomers • Alpha satellite DNA • CENP- B
Chromosomes as functioning organelles Telomeres Telomeres are specialized structures, comprising DNA and protein, which cap the ends of eukaryotic chromosomes. Functions • Maintaining the structural integrity of a chromosome. • Ensuring complete replication of the extreme ends of chromosomes. • Helping establish the three-dimensional architecture of the nucleus and/or chromosome pairing. The ability of telomerase to replicate a chromosome end depends on the unique molecular structure of the telomere.
Chromosomes as functioning organelles Telomeres • Eukaryotic telomeres consist of a long array of tandem repeats. One DNA strand contains TG-rich sequences and terminates in the 3′ end; the complementary strand is CA-rich. • Highly conserved in evolution - there is considerable similarity in the simple sequence repeat, • Example • TTGGGG (Paramecium), TAGGG (Trypanosoma) TTTAGGG (Arabidopsis) and TTAGGG (Homo sapiens)
Chromosomes as functioning organelles Telomeres • First isolated from the protozoan Tetrahymena thermophila and possess multiple copies of the sequence:
Human Telomeres • The (TTAGGG)" array of a human telomere spans about 10-15 kb. • A very large protein complex shelterin, or the telosome contains several components that recognize and bind to telomeric DNA. • Two telomere repeat binding factors (TRFl and TRF2) bind to double-stranded TTAGGG sequences. • G-rich strand has a Single-stranded overhang at its 3' end that is typically 150-200 nucleotides long. • This can fold back and form base pairs with the other, C-rich, strand to form a telomeric loop known as theT-loop. Protect the telomere DNA from natural cellular mechanisms that repair double-stranded DNA breaks.
Chromosomes as functioning organelles Telomeres • Telomeres have now been isolated from protozoans, plants, humans, and other organisms; most are similar in structure.
Chromosomes as functioning organelles Telomere Structure • The G-rich strand often protrudes beyond the complementary C-rich strand at the end of the chromosome. • The length of the telomeric sequence varies from chromosome to chromosome and from cell to cell, suggesting that each telomere is a dynamic structure that actively grows and shrinks. • The telomeres of Drosophila chromosomes are different in structure. • They consist of multiple copies of the two different retrotransposons , Het-A and Tart,arranged in tandem repeats. • Apparently, in Drosophila, loss of telomere sequences during replication is balanced by transposition of additional copies of the Het-A and Tart elements.
Telomerase and the chromosome end-replication problem • Telomerase is a ribonucleoprotein enzyme whose polymerase function is criticaliy dependent on an RNA subunit, TERC (telomerase RNA component), and a protein subunit, TERT (telomerase reverse transcriptase). • At the 5‘ end of vertebrate TERC RNA is a hexanucleotide sequence that is complementary to the telomere repeat sequence. • It will act as a template to prime the extended DNA synthesis of telomeric DNA sequences on the leading strand. • Further extension of the leading strand provides the necessary template for DNA polymerase to complete the synthesis of the lagging strand.
Chromosomes as functioning organelles: origins of replication Origins of replication are the sites where DNA synthesis begins; • Eukaryotic chromosomes have multiple origin of replication. • Eukaryotic origins of replication; yeast, where the presence of a putative replication origin can be tested by a genetic assay. • Bacterial replication origin in the plasmid does not function in yeast, therefore the few plasmids that transform at high efficiency must possess a sequence within the inserted yeast fragment that confers the ability to replicate extrachromosomally at high efficiency - that is an autonomously replicating sequence (ARS) element.
Chromosomes as functioning organelles: origins of replication • ARS elements are thought to derive from authentic origins of replication and, in some cases, this has been confirmed by mapping a specific ARS element to a specific chromosomal location and demonstrating that DNA replication is indeed initiated at this location. • ARS elements extend for only about 50 bp and consist of an AT-rich region which contains a conserved core consensus and some imperfect copies of this sequence. • ARS elements contain a binding site for a transcription factor and a multiprotein complex is known to bind to the origin.
Chromosomes as functioning organelles: origins of replication • Mammalian replication origins are less well defined because of the absence of a genetic assay. • There are speculations that replication can be initiated at multiple sites over regions tens of Kb long. • Computer analysis of regions encompassing several eukaryotic origins of replication, including some human and other mammalian examples, identified a consensus DNA sequence WAWTTDDWWWDHWGWHMAWTT where W = A or T; D = A or G or T; H = A or C or T; and M = A or C
Cytogenetics • Is the study of the structure and properties of chromosomes, chromosomal behaviour during mitosis and meiosis, chromosomal influence on the phenotype and the factors that cause chromosomal changes. • Related to disease status caused by abnormal chromosome number and/or structure.
Methods for chromosomal analysis: Karyotyping and banding The collection of all the chromosomes is referred to as a Karyotype. The method used to analyze the chromosome constitution of an individual, known as chromosome banding. Chromosomes are displayed as a karyogram.
Obtaining and preparing cells for chromosome analysis • Cell source: • Blood cells • Skin fibroblasts • Amniotic cells / chorionic villi • Increasing the mitotic index - proportion of cells in mitosis using colcemid • Synchronizing cells to analyze prometaphase chromosomes
Key procedure • In the case of peripheral (venous) blood • A sample is added to a small volume of nutrient medium containing phytoheamagglutinin, which stimulates T lymphocytes to divide. • The cells are cultured under sterile conditions at 37C for about 3 days, during which they divide, and colchicineis then added to each culture. • This drug has the extremely useful property of preventing formation of the spindle, thereby arresting cell division during metaphase, the time when the chromosomes are maximally condensed and therefore most visible. • Hypotonic saline is then added, which causes the red blood cells to lyze and results in spreading of the chromosomes, which are then fixed , mounted on a slide and stained ready for analysis
Karyotype Analysis • Following Steps are involved; • Counting the number of cells, sometimes referred as metaphase spread • Analysis of the banding pattern of each individual chromosome in selected cells. • Total chr. Count is determined in 10-15 cells, but if mosaicism is suspected then 30 or more cell count will be undertaken. • Detailed analysis of the banding pattern of the individual chromosomes is carried out in approx. 3-5 metaphase spread, which shows high quality banding. • The banding pattern of each chromosome is specific and shown in the form of Idiogram.