Download
1 / 8
90 likes | 194 Views
Explore the intricate pathways and enzymes involved in starch metabolism from algae to plants, shedding light on enzymatic actions and metabolic steps. Discover the conservation of key steps and their biological significance.

E N D
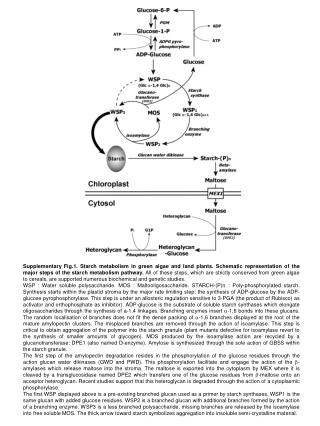
Supplementary Fig.1. Starch metabolism in green algae and land plants. Schematic representation of the major steps of the starch metabolism pathway. All of these steps, which are strictly conserved from green algae to cereals, are supported numerous biochemical and genetic studies. WSP : Water soluble polysaccharide. MOS : Maltooligosaccharide. STARCH-(P)n : Poly-phosphorylated starch. Synthesis starts within the plastid stroma by the major rate limiting step: the synthesis of ADP-glucose by the ADP-glucose pyrophosphorylase. This step is under an allosteric regulation sensitive to 3-PGA (the product of Rubisco) as activator and orthophosphate as inhibitor). ADP-glucose is the substrate of soluble starch synthases which elongate oligosaccharides through the synthesis of a-1,4 linkages. Branching enzymes insert a-1,6 bonds into these glucans. The random localisation of branches does not fit the dense packing of a-1,6 branches displayed at the root of the mature amylopectin clusters. The misplaced branches are removed through the action of isoamylase. This step is critical to obtain aggregation of the polymer into the starch granule (plant mutants defective for isoamylase revert to the synthesis of smaller amounts of glycogen). MOS produced by the isoamylase action are recycled by a glucanotransferase: DPE1 (also named D-enzyme). Amylose is synthesized through the sole action of GBSS within the starch granule. The first step of the amylopectin degradation resides in the phosphorylation of the glucose residues through the action glucan water dikinases (GWD and PWD). This phosphorylation facilitate and engage the action of the b-amylases which release maltose into the stroma. The maltose is exported into the cytoplasm by MEX where it is cleaved by a transglucosidase named DPE2 which transfers one of the glucose residues from b-maltose onto an acceptor heteroglycan. Recent studies support that this heteroglycan is degraded through the action of a cytoplasmic phosphorylase. The first WSP displayed above is a pre-existing branched glucan used as a primer by starch synthases. WSP1 is the same glucan with added glucose residues. WSP2 is a branched glucan with additional branches formed by the action of a branching enzyme. WSP3 is a less branched polysaccharide, missing branches are released by the isoamylase into free soluble MOS. The thick arrow toward starch symbolizes aggregation into insoluble semi-crystalline material.
E. histolytica A. tumefaciens N. punctiforme Cyanothece CCY0110 P. marinus 25 S. elongatus N. crassa A. fumigatus 22 S. cerevisiae 1 G. gallus 1 X. tropicalis 1 99 M. musculus 1 H. sapiens 1 V. carteri 1 91 C. reinhardtii 1 O. tauri M. pusila RCC299 92 M. pusila CCMP1545 O. sativa ch A. thaliana ch S. tuberosum ch P. sativum ch P. trichocarpa ch D. dictyostelium A. thaliana cy2 44 A. thaliana cy1 O. sativa cy 99 P. trichocarpa cy S. tuberosum cy P. sativum cy V. cholerae E. coli 100 Y. pestis S. usitatus M. gilvum Synechocystis sp. PCC6803 91 T. vaginalis 17 E. histlytica A. fumigatus 2 S. cerevisiae 2 100 V. carteri 2 C. reinhardtii 2 T. cruci X. tropicalis 2 38 G. gallus 2 0.5 substitution per site M. musculs 2 H. sapiens 2 Supplementary Fig.2. Plant phosphoglucomutases were duplicated from a single eukaryotic gene. Maximum likelihood unrooted tree inferred for phosphoglucomutases enzymes. The scalebar represent the branch length corresponding to 1 substitution per site. Bootstrap values are indicated at corresponding nodes. Three subgroups can be clearly observed. Two subgroups at the bottom of the tree are a bacterial subgroup and a eukaryotic subgroup. Two isoforms of Chlamydomonas and Volvox can be observed in this latter. Enzymes form which involvement in starch meatbolism is experimentaly proven are all related within the upper subgroup. Inferring the tree with only these sequences support the Eukaryotic origin of the plant phosphoglucomutases.
V. carteri sBE2b C. reinhardtii sBE2b V. carteri sBE2a 56 C. reinhardtii sBE2a O. lucimarinus sBE2 O. tauri sBE2 88 M. pusilla CCMP1545 sBE2 M. pusilla RCC299 sBE2 A. thaliana sBE2a 97 A. thaliana sBE2b S. tuberosum sBE2 P. trichocarpa sBE2 46 O. sativa sBE2b O. sativa sBE2a O. sativa sBE1 P. trichocarpa sBE1 S. tuberosum sBE1 76 O. lucimarinus sBE1 56 O. tauri sBE1 M. pusilla RCC299 sBE1 M. pusilla CCMP1545 sBE1 C. reinhardtii sBE1 V. carteri sBE1 H. sapiens S. cerevisiae A. fumigatus 59 C. elegans 0.5 substitution per site D. melanogaster 100 M. pusilla RCC299 sBE3 100 M. pusilla CCMP1545 sBE3 B. subtilis G. violaceus P. marinus 77 Synechocystis Cyanothece CCY0110 1 C. watsonii 1 C. watsonii 2 100 Cyanothece CCY0110 2 A. tumefaciens E. coli Y. pestis 33 Cyanothece CCY0110 3 C. wasonii 3 P. trichocarpa sBE3 A. thaliana sBE3 Supplementary Fig.3. Plant branching enzymes are of eukaryotic origin. Maximum likelihood unrooted tree inferred for starch branching enzymes. Bootstrap values are indicated at corresponding nodes. Every plant and green alga contain one type I and at least one type 2 branching enzymes. The existence of a bacteria-related branching enzyme in Populus and Arabidopsis was already reported (Han et al. 2007), this enzyme is probably not involved in starch metabolism (Dumez et al. 2005). A third subgroup of undefined phylogeny can be distinguished in Micromonas species. The scalebar represent the branch length corresponding to 0.5 substitution per site.
C. reinhardtii V. carteri 58 M. pusilla CCMP1545 M. pusilla RCC299 D-enzymes 93 O. lucimarinus O. tauri O. sativa 99 S. tuberosum P. trichocarpa A. thaliana A. variabilis Synechocystis Cyanobacterial glucanotransferases C. watsonii G. variabilis P. marinus 41 O. sativa A. thaliana P. trichocarpa S. tuberosum 68 O. tauri O. lucimarinus M. pusilla RCC299 Transglucosidases M. pusilla CCMP1545 100 V. carteri C. reinhardtii D. discoideum 0.5 substitution per site E. histolytica 1 26 E. histolytica 2 T. vaginalis 1 T. vaginalis 2 T. vaginalis 3 Supplementary Fig.4. Two kind of alpha-glucanotransferase are detected in every plants. Maximum likelihood unrooted tree inferred for alpha-1,4-glucanotransferases. Bootstrap values are indicated at corresponding nodes. This tree supports the phylogenetic divergence between the two kinds of glucanotransferase activities. D-enzymes (DPE1) are clearly related to bacterial glucanotransferase (which catalyse the same reaction) while transglucosidases (DPE2) seem only to exist in eukaryotic cells. The scalebar represent the branch length corresponding to 0.5 substitution per site.
Supplementary Table 1. Complete references of the annotated genes used in this work. These sequences can be retrieved using the gene name as search criteria on the JGI website pages (http://genome.jgi-psf.org/) of each genome. PART 1
Micromonas RCC299 CCMP1545 Phosphoglucomutase - e_gw2.01.168.1 estExt_fgenesh1_kg.C_20035 ADPG- Small estExt_fgenesh2_kg.C_Chr_070019 estExt_fgenesh1_kg.C_10041 Pyrophosphorylase e_gw2.01.322.1 estExt_Genewise1Plus.C_20473 Large estExt_Genewise2Plus.C_Chr_020427 e_gw1.4.863.1 Starch synthases gbss estExt_fgenesh2_kg.C_Chr_020003 estExt_fgenesh1_kg.C_110003 estExt_fgenesh2_kg.C_Chr_060061 estExt_fgenesh1_pg.C_50151 SSI gw2.11.73.1 gw1.8.77.1 e_gw2.15.329.1 estExt_Genewise1.C_140367 SSII e_gw2.07.522.1 fgenesh1_pm.C_scaffold_9000005 estExt_fgenesh2_kg.C_Chr_150005 estExt_Genewise1Plus.C_50295 SSIII estExt_fgenesh2_kg.C_Chr_060062 estExt_fgenesh1_pg.C_140161 e_gw2.08.94.1 e_gw1.11.141.1 SSIV estExt_fgenesh2_kg.C_Chr_030028 estExt_fgenesh1_pm.C_170017 SSV - - Branching enzymes SBE1 e_gw2.08.666.1 e_gw1.8.541.1 SBE2 estExt_Genewise2Plus.C_Chr_010412 estExt_fgenesh1_kg.C_20047 "SBE3" e_gw2.08.666.1 fgenesh1_pm.C_scaffold_5000096 Isoamylases Isa1 estExt_fgenesh2_pg.C_Chr_130076 estExt_Genewise1Plus.C_70118 Isa2 EuGene.0100010130 est_orfs.2_3198_4270363:1 Isa3 estExt_fgenesh2_kg.C_Chr_150010 estExt_fgenesh1_pm.C_140015 Pullulanase - fgenesh2_pm.C_Chr_07000017 estExt_Genewise1.C_10891 Glucano-Transferases D-enz estExt_Genewise2.C_Chr_010539 estExt_Genewise1.C_20623 DPE2 e_gw2.12.29.1 e_gw1.10.23.1 estExt_fgenesh2_kg.C_Chr_110012 estExt_fgenesh1_kg.C_80004 Phosphorylases estExt_fgenesh2_kg.C_Chr_120004 estExt_Genewise1Plus.C_100032 e_gw2.12.509.1 estExt_Genewise1Plus.C_100176 e_gw2.04.10.1 estExt_fgenesh1_pg.C_50208 Glucan dikinases GWD fgenesh2_kg.C_Chr_06000057 estExt_fgenesh1_pg.C_90205 e_gw2.12.513.1 estExt_fgenesh1_pg.C_100126 est_orfs.6_8624_4273671:1 estExt_fgenesh2_kg.C_Chr_050017 e_gw1.9.75.1 PWG e_gw2.09.6.1 EuGene.0000050241 estExt_fgenesh2_kg.C_Chr_060065 EuGene.0000050242 est_cluster_kg.Chr_04__16__3199517:1 e_gw1.12.72.1 Beta-amylases - fgenesh2_kg.C_Chr_11000018 e_gw1.8.113.1 fgenesh2_kg.C_Chr_14000003 fgenesh2_kg.C_Chr_02000034 gw1.12.147.1 e_gw2.02.334.1 estExt_Genewise1.C_40491 fgenesh2_kg.C_Chr_06000003 Alpha-amylases - estExt_Genewise1.C_40669 fgenesh2_pm.C_Chr_09000077 estExt_Genewise1.C_90048 EuGene.0600010408 fgenesh1_pg.C_scaffold_18000135 EuGene.1100010582 gw2.15.105.1 Mex1 - estExt_fgenesh2_kg.C_Chr_140010 EuGene.0000030121 Sex4 - gw2.05.501.1 estExt_fgenesh1_pg.C_30481 Table 1. PART 3
ESTs Transit peptide Micromonas Ostreococcus Chlamydomonas Volvox CCMP 1545 RCC299 tauri lucimarinus Phosphoglucomutase 1 ~ 50 Y N Y N N N Phosphoglucomutase 2 - Y N n/a n/a n/a n/a ADP-glucose pyrophosphorylase S 2 Y Y Y N N N ADP-glucose pyrophosphorylase L1 ~ 40 N Y Y N N N ADP-glucose pyrophosphorylase L2 1 N N Y N N N Granule bound starch synthase 1 ~ 60 Y Y Y N N N Granule bound starch synthase 2 - N n/a n/a n/a n/a n/a Soluble starch synthase 1a 1 Y N N N N N Soluble starch synthase 1b - N N Y N n/a n/a Soluble starch synthase 2a ~ 20 N Y N N N N Soluble starch synthase 2b n/a n/a n/a Y Y n/a n/a Soluble starch synthase 3a ~ 15 N N N N N N Soluble starch synthase 3b - N Y N N N Y Soluble starch synthase 3c n/a n/a n/a N N N N Soluble starch synthase 4 ~ 30 Y Y N N n/a N Soluble starch synthase 5 8 N N n/a n/a n/a n/a Branching enzyme 1 - N N N Y Y N Branching enzyme 2a 4 N N Y N N N Branching enzyme 2b ~ 50 N N n/a n/a n/a n/a Branching enzyme 3 3 Y Y Y N n/a n/a Isoamylase 1 2 Y N N N Y Y Isoamylase 2 - Y N N Y N N Isoamylase 3 ~ 15 Y N N Y N N Pullulanase - N N N Y N Y D-enzyme ~ 10 Y N Y N Y N Transglucosidase - N N N N N N Phosphorylase 1 ~ 50 N Y Y N Y N Phosphorylase 2 ~ 20 N Y Y N N N Phosphorylase 3 n/a n/a n/a N N N N Glucan water dikinase 1 ~ 10 N N N N N N Glucan water dikinase 2 ~ 10 N N N N Y N Glucan water dikinase 3 1 N n/a Y N N N Phosphoglucan water dikinase 1 1 Y N N N N N Phosphoglucan water dikinase 2 n/a n/a N N N N N Phosphoglucan water dikinase 3 n/a n/a n/a Y Y n/a n/a Phosphoglucan water dikinase 4 n/a n/a n/a Y n/a n/a n/a b -amylase 1 ~ 40 N N N N N N b -amylase 2 2 N N N N N N b -amylase 3 10 N n/a n/a n/a n/a n/a a -amylase 1 ~ 15 N Y N N N N a -amylase 2 ~ 20 N N N N N N a -amylase 3 ~ 10 Y n/a N N N N a -amylase 4 n/a n/a n/a N N n/a N a -amylase 5 n/a n/a n/a N N n/a n/a a -amylase 6 n/a n/a n/a n/a N n/a n/a a -amylase 7 n/a n/a n/a n/a N n/a n/a a -amylase 8 n/a n/a n/a n/a N n/a n/a Maltose exporter 4 Y N N N N N Sex4 10 Y N N N N N In the first column, under the heading "ESTs", we have listed numbers that correspond to the number of different ESTs found in the extensive databases generated for Chlamydomonas exclusively. In the other columns, we have symbolise by "Y" and "N" respectively the presence or abscence of a putative transit peptide as detected using ChloroP (http://www.cbs.dtu.dk/services/ChloroP/). We insist that this detection is meaningless as many plastidials proteins detected by proteomics lack transit peptides and because some gene that contain very clear targeting sequences may not at all be located in plastids. A good example in starch metabolism is afforded by the Arabidopsis DPE2 protein wich contains a transit peptide and yet is exclusively located in the cytosol. Supplementary Table 2. Transit peptide predictions and EST survey.