Download

1 / 1
10 likes | 185 Views
Standard Screen. Salt Screen. #. #. Hits. Hits. Precipitant. Precipitant. Salt. Salt. Buffer. Buffer. Like*. Like*. 75. 73. 11. 8. 30%(w/v) PEG 8000. 30%(w/v) PEG 4000. 0.2 M (NH 4 ) 2 SO 4. 0.2 M Li 2 SO 4. 0.1 M Cacodylate pH 6.5. 0.1 M Tris pH 8.5. H1 #15. H1 #17. 75.

E N D
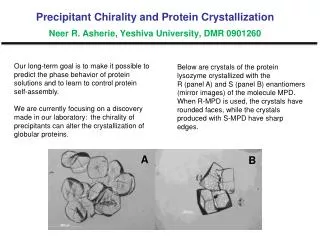
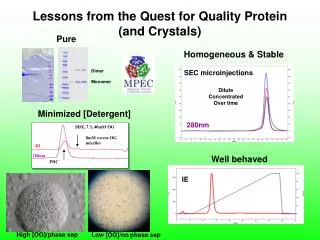
Standard Screen Salt Screen # # Hits Hits Precipitant Precipitant Salt Salt Buffer Buffer Like* Like* 75 73 11 8 30%(w/v) PEG 8000 30%(w/v) PEG 4000 0.2 M (NH4)2SO4 0.2 M Li2SO4 0.1 M Cacodylate pH 6.5 0.1 M Tris pH 8.5 H1 #15 H1 #17 75 41 7 6 30%(w/v) PEG 8000 30%(w/v) PEG 4000 0.2 M Li2SO4 0.2 M Li2SO4 0.1 M Tris pH 8.5 0.1 M NaAcetate pH 4.5 H1 #17 W1 #17 76 85 5 6 20%(w/v) PEG 8000 2.0 M (NH4)2SO4 0.2 M MgAcetate 0.1 M Cacodylate pH 6.5 H1 #32 H1 #18 89 70 5 5 30%(w/v) PEG 4000 1.4 M Na Citrate 0.2 M (NH4)2SO4 0.1 M HEPES pH 7.5 0.1 M NaCitrate pH 5.6 H1 #38 H2 #9 43 70 5 4 30%(w/v) PEG 4000 20%(w/v) PEG 8000 0.2 M MgCl2 0.2 M (NH4)2SO4 0.1 M NaCitrate pH 5.6 0.1 M Tris pH 8.5 H2 #9 W2 #2 2 5 20%(w/v) PEG 3000 0.1 M NaCitrate pH 5.5 W1 #6 Target Residue Ser Thr His Tyr Overlaps* Overlaps* Overlaps* Overlaps* Standard Salt Standard Salt Standard Salt Standard Salt Standard Salt Total Unique° Total Unique° Mutant Distribution of Hits Standard Screen * Overlaps are conditions that produced hits in both types of screens. ° Unique refers to how many different conditions produced hits for the mutant series The Best Conditions Distribution of Hits Salt Screen *H= Hampton Research Crystal Screen W=Emerald Biostructures Wizard Screen DY2 K138Y, K141Y - A second from of DY 30% PEG 6000, 0.1M bicine pH=7.5 Resolution 2.2Å Spacegroup P21 (a=39.5,b=56.2,c=61.6,=91.7) Discontinued: Solubility Problems A B C D CT K135T, K138T, K141T E F G H I 35% PEG 4K, 0.2M Li2SO4, 0.1M Tris pH 8.5 Resolution 2.0 Å Preparation of High Quality Protein Crystals by Surface Entropy Reduction - Tyrosine as a Crystal Contact Catalyst. Tomek Boczek, David R. Cooper, Kasia Grelewska, Gosia Pinkowska, Gosia Sikorska, Michal Zawadzki, and Zygmunt DerewendaPSI Center for Structure and Function Innovation and Department of Molecular Physiology and Biological Physics University of Virginia. Charlottesville, VA, 22908 The Results The Screens Abstract Crystallization is a limiting step in macromolecular crystallography. In cases where proteins are particularly recalcitrant to crystallization efforts, mutational modification of surface properties may be essential. We previously suggested that targeted replacement of clusters of residues with high-conformational entropy (lysines, glutamates and/or glutamines) with alanines leads to formation of epitopes that are capable of mediating crystal contacts. This is because the entropic cost of immobilizing large side chains at the intermolecular contact regions has been reduced and crystal contacts can be formed by the mutated epitopes. This Surface Entropy Reduction (SER) method has facilitated the crystallization and structure determination of a number of novel proteins and has also led to the discovery of crystal forms that diffract to significantly higher resolution than the wild-type form. However, it has not been conclusively demonstrated if alanine constitutes the best choice for replacement of high-entropy residues. Here we present a systematic study of the replacement of nine Lys/Glu-rich patches in RhoGDI with four target residues; Ser, Thr, His and Tyr. All four amino acids are known to occur at interfaces with significantly higher incidence than Lys or Glu / Gln, and may mediate weak protein-protein interactions leading to crystal formation. Our results show that tyrosine is a particularly good choice for the target amino acid, with threonines and histidines also performing quite well. The mutated residues often participate in crystal contacts, in both homotypic (symmetric) or heterotypic (head-to-tail) intermolecular interactions. We also examined a crystallization method proposed by Janet Newman that replaces the normal crystallization reservoir solutions with 1.5 M NaCl. The results are very promising; with more than half of the mutants in this series yielding more crystals when salt was used as the reservoir solution. Moreover, this method greatly increased the variety of conditions that yielded crystals, with little overlap of the conditions that yielded crystals for the two types of screens. This suggests a crystallization strategy for proteins for which crystallization is the major bottleneck. Creating several mutants by replacing patches glutamates and lysines with two or more target residues and conducting screens with normal and alternate reservoir solutions greatly increases the chances of obtaining diffraction quality crystals • The Super Screen • We use a custom 96 well screen we had Nextal generate for us. • For details see, • http://ginsberg.med.virginia.edu/~dcoop/Superscreen • We use sitting drop setups. For all of our screens, the protein was concentrated to 15 mg/ml. • Standard Screens • Drops are mixed 1:1 with protein solution and Super Screen reagent. • Reservoir is 100 l of Super Screen reagent. • Salt Screens • As above, drops are mixed 1:1 with protein solution and Super Screen reagent. • Reservoir is 100 l of 1.5 M NaCl • This has the advantage of significantly reducing the cost of crystallization setups. 10 ml screen reagent = 10,000 drops. • Things To Note • Standard Screens • Tyrosine, by far, scored the most hits. Three times as many as the next highest, threonine. • Almost half of tyrosine’s hits were for the DY mutant, and more than half of threonine’s hits were the IT mutant. • Histidines were the most consistent, with 7 of 8 mutants yielding hits. • Salt Screens • Once again, tyrosine produces the most hits. • In 20 of 32 mutants, the salt screen produced as many or more hits than the standard screen. • Threonines were the most consistent, with 7 of 8 mutants yielding hits. • The two screens yielded crystals in different conditions. There are very few conditions that produce hits in both screens. • Overall • Of the 16 screens performed for each target residue, threonine and histidine had the most screens (12) yielding crystals. Tyrosine had 11. • Threonine was the only target residue that produced a hit for every mutant when both screens were performed. The other target residues yielded crystals for 7 of 8 mutants. • There seems to be a preference for a particular target residue for some mutants. The D mutant worked best with tyrosine and the I mutant worked best with threonine. • For each mutant, 3 of the 4 target residues yielded crystals when both screens were performed. Therefore, if we had used two target residues with any one mutant, we would have found at least one crystal producing condition. The Crystals Experimental Design Old and New RhoGDI Crystal Forms Previous Xtals Model Protein : RhoGDI (66-204) RhoGDI posses all of the characteristics of proteins that lend themselves to crystallization by the SER method. It expresses and purifies well and can be concentrated easily, but wild-type RhoGDI is difficult to crystallize. Additionally RhoGDI is rich in lysines (10.1% -- average frequency is 7.2% ) and glutamates (7.9% -- average frequency is 3.7%), giving us many potential mutation sites. New Xtals References DY K138Y, K141Y • Janet Newman • Rebecca Page • Other Crystal Screen Paper • Zygmunt’s Methods • Plug Luki The Mutant Series Nine mutants were chosen, with the mutations designed to reduce or eliminate clusters of high-entropy residues. The mutants were designated by two letters; the first letter indicates which mutations the mutant contains and the second letter designates the target amino acid. Thus, the CY mutant is K135Y, K138Y, K141Y. The B mutant was discontinued do to low expression or solubility. 32% PEG 8000, 0.22M (NH4)2SO4, 0.1M Cacodylate pH=6.5 Resolution 2.1Å Spacegroup P21 (a=32.0,b=55.1,c=38.9,=107.5) Conclusions CY K135Y, K138Y, K141Y We have previously shown that replacing large, highly-entropic residues with alanines can facilitate crystallization. The experiment described here demonstrates that other residues can be used to replace surface exposed lysines and glutamates. This study reveals that tyrosines, threonines and histidines are suitable target amino acids for crystallization by the SER method. Overall, tyrosines produced the most impressive statistics for “number of hits”, but threonines and histidines appear to be slightly more consistent. In several cases, one target residue produced many more hits for a particular mutant than did the other target residues. This mutant / target residue correlation leads us to conclude that a wise strategy for crystallization by the SER method would be to choose a cluster of highly entropic residues to mutate, and mutate them to two or more target residues. Additionally, this experiment examined a recent approach to crystal screening that uses an alternate reservoir solution, instead of the traditional mother liquor. The alternate reservoir screens produced even more hits than the traditional screening method. Additionally, this type of screening gave rise to crystals in different conditions than the standard screen did. The efficacy and efficiency of this type of screening makes it a tool that any crystallographer should employ. All RhoGDI mutants were expressed as a GST-fusion protein. After cleavage of the fusion protein with rTEV, GST was removed by size exclusion chromatography. GST-RhoGDI 4.0 M Sodium Formate Resolution 2.50Å Spacegroup C21 (a=69.2,b=78.0,c=50.9,=91.5) RhoGDI Initial Purification Size Exclusion Purification