Download

1 / 63
780 likes | 1.56k Views
Heat Shock Proteins, Hsp90 Inhibitors, and Protein Degradation. By: Vince Centioni Paper: “A high affinity conformation of Hsp90 confers tumour selectivity on Hsp90 inhibitors”. I. Background A. Protein Degradation.

E N D
Heat Shock Proteins, Hsp90 Inhibitors, and Protein Degradation By: Vince Centioni Paper: “A high affinity conformation of Hsp90 confers tumour selectivity on Hsp90 inhibitors”
I. BackgroundA. Protein Degradation • Protein molecules are continuously synthesized and degraded in all living organisms. The concentration of individual cellular proteins is determined by a balance between the rates of synthesis and degradation, which in turn are controlled by a series of regulated biochemical mechanisms. • The only way that cells can reduce the steady state level of a particular protein is by degradation. Thus, complex and highly-regulated mechanisms have been evolved to accomplish this degradation.
A. Protein Degradation cont. • General principles: Peptide bond: Planar amide bond between α-carboxyl group & α-amino group of 2 adjacent amino acids. • Proteolysis: Biochemical degradation of protein through hydrolysis of peptide bonds.
A. General Principle Diagram: Peptide + Water Acid + Amine
A. Protein Degradation cont. (Intracellular Proteolytic Systems) • Lysosomal and Non-lysosomal • Lysosomal: Steps= Uptake (Autophagy) into lysosome: Secretory vesicles; Cytoplasm; Organelles, and enzymatic degradation . • Non-lysosomal: Steps= Tagging of protein to be degraded (by ubiquitnation). Recognition of proteolytic system: exposure of peptide sequence or distinction of unfolded protein segments • Example : Proteasome
A. Protein Degradation cont. • Cells which are subject to stress such as starvation, heat-shock, chemical insult or mutation respond by increasing the rates of degradation. • Selective degradationof particular proteins may occur in response to internal and external signals.
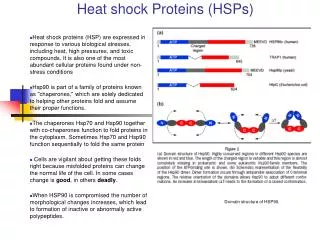
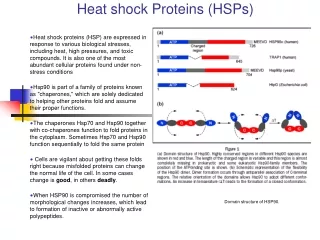
B. Heat Shock Proteins • Heat Shock proteins: Family of proteins found in all cells that are expressed in response to cold, heat and other environmental stresses. • Some serve to stabilize proteins in abnormal configurations, and play a role in folding and unfolding of proteins, acting as molecular chaperones. • There are four major subclasses: Hsp90, Hsp70, Hsp60, and small Hsps
B. Heat Shock Proteins cont. • Heat shock proteins are induced when a cell undergoes various types of environmental stresses like heat, cold, and oxygen deprivation. • Heat shock proteins are also present in perfectly normal conditions where they act as chaperones making sure that the cell’s proteins are in the right shape and in the right place at the right time. • HSPs also help shuttle proteins from one compartment to another inside the cell, and transport old proteins to “garbage disposals” inside the cell.
B. Heat Shock Proteins cont. • Heat shock proteins and the immune system: under normal conditions HSPs are found outside the cell. But if a cancerous or infected cell has become so sick that it dies and its membrane bursts, all of cell’s contents spill out, including Hsps that are bound to peptides. • These extracellular Hsps send a very strong “danger r signal” to the immune system, instructing it to destroy the other diseased cells.
C. Hsp-danger signal to the immune system: • A sick cell dies and ruptures, spilling the Hsp-peptide complexes. • These extracellular complexes of HSPs and peptides are detected by circulating immune system cells called APCs (antigen-presenting cells). • The Hsp complexes bind the CD91 receptor on the APC cell surface. The APC can then take in the Hsp complexes, and then they travel to the lymph nodes.
C. Hsp-danger signal cont. • In the lymph nodes, the APCs take the peptides that were associated with HSPs and re-represent them on the cell surface. • Specialized immune cells called T cells “see” these peptides and are then programmed to seek out the cells bearing these specific, abnormal peptides. • Because every person and every cancer is different, the unique repertoire of antigenic peptides represents that individual specific cancer’s “fingerprint.”
D. Heat Shock Protein 90 • Cellular chaperone protein required for the activation of several eukaryotic protein kinases, including the cyclin-dependant kinase (CDK4). • Geldanamycin, and inhibitor of the protein-refolding activity of Hsp90, has been shown to have antitumor activities. • Hsp90 is the most abundant heat shock protein under normal conditions.
D. Hsp90 cont. • The protein exists as two major isoforms, Hsp90-alpha and Hsp90-beta. • The protein’s activity has been shown to be ATP dependant with a unique pocket located in the N-terminal region. • This pocket is the site where ATP and ADP binding activity take place. Once ATP binds , a structural amendment by Hsp90 induces a conformational change from an open position to a closed position.
D. Hsp90 cont. • Hsp90’s contributions in signal transduction, protein folding, and protein degradation. • Hsp90 is a phosphoprotein containing two or three covalently bound phosphate molecules per monomer, and the phosphorylation is thought to enhance its function. The monomer of the Hsp90 consists of a conserved 25-kaDa N-terminal domain and a 55-kaDa C-terminal domain linked by a 35-kaDa charged linker region. • Together the C-terminal domain and the linker region helps in the dimerization of the protein
D. Hsp90 cont. • Hsp90 exhibits ATPase activity which is essential for its chaperone function. • Hsp90 binds to an array of client proteins, where its co-chaperone specificity varies and depends on the actual client protein. • There is a growing list of Hsp90 client proteins and most of them include molecules involved in signal transduction.
D. Hsp90 cont. • Hsp90 forms several discrete sub-complexes, each containing different set of co-chaperones that function at different steps during the folding process of the client protein. • Unlike other chaperones, Hsp90 contains two independent chaperone sites that differ in their substrate specificity, probably working in the form of a switch between Hsp90 and Hsp70 client protein interactions.
D. Hsp90 cont. (Multi-chaperone complex) • The best understood molecular association of Hsp90-multi-chaperone complexes was in conjunction with the maturation of steroid receptors. • The folding process of steroid receptors and their translocation to the cell nucleus requires Hsp90. • Steroid receptors also require molecular chaperones for their ligand binding, transcriptional activation and repression after stimulus.
D. Hsp90 cont. • Though there are reports that some Hsp90 co-chaperones can work independently of Hsp90, the full competence of these co-chaperones requires Hsp90. • Co-chaperones also help Hsp90-client protein binding interactions between Hsp70 client proteins, and docking of cytoskeletal proteins.
III. IntroductionA. Hsp90 and Cancer Cells • Hsp90 has been implicated in the survival of cancer cells. • Hsp90 regulates the function and stability of many key signal proteins that help cancer cells to escape the inherent toxicity of their own environment, to evade chemotherapy, and to protect themselves from the results of their own genetic instability.
A. Hsp90 and Cancer Cells cont. • Hsp90 plays a key role in the conformational maturation of oncogenic signaling proteins, including HER-2/ErbB2, Akt, Raf-t, Bcr-Abl and mutated p53. • Tumor Hsp90 is present entirely in multi-chaperone complexes with high ATPase activity. • Tumor cells overexpress Hsp90 client proteins, suggesting that a greater amount of Hsp90 in tumor cells might be engaged in active chaperoning and present in multichaperone complexes that could modulate the binding affinity of ligands to Hsp90.
A. Hsp90 and Cancer Cells cont. • Hsp90 regulates many signaling pathways in cancer cells. • Recent discoveries in cell biology have demonstrated that many of the key signaling molecules that are deregulated in human cancers require the action of Hsp90 chaperone family in order to maintain their function.(Neckers and Lee 2003)
A. Hsp90 and Cancer Cells cont. • Cancer cells have numerous abnormalities that make them more dependent upon growth and survival which, are, in turn dependent upon Hsp90. • Hsp90 works with other chaperone proteins and forms a Hsp90 multichaperone complex that maintains tumor progression, by stabilizing and interacting with a growing list of various kinases. • Hsp90 chaperone complexes control protein folding and influence protein degradation.
A. Hsp90 multichaperone pathway and Ubiquitin-mediated degradation pathway:
A. Hsp90 and Cancer Cells cont. • In tumors these signaling proteins are deregulated resulting in uncontrolled cell growth and survival. • Certain Hsp90 inhibitors are designed to eliminate these deregulated signal transduction molecules from the tumor cell leading to death of the tumor.
B. Hsp90 Inhibitors • Certain Hsp90 inhibitors can bind into the ATP binding site of Hsp90 altering the function of the Hsp90 multichaperone complex. • These inhibitors convert the Hsp90 complex from a catalyst for protein folding into one that induces protein degradation. • The result is the degradation of a specific set of cancer signaling molecules, leading to cell cycle arrest and tumor cell death.
B. Hsp90 inhibitors cont. • Currently used Hsp inhibitors include geldanaymicn, herbimycin A and 17-AAG. • In 1994, certain ansamycins were found to bind to Hsp90 and to cause the degradation of client proteins including Src kinases. • Further efforts to develop anticancer drugs were made using geldanamycin analogs, and 17AAG was chosen as the best candidate for clinical trials.
B. Hsp90 Inhibitors cont. • Geldanaymicn: is a natural Hsp90 inhibitor, that essentially causes the complete destruction of the receptor tyrosine kinase HER2/neu, a key driver of breast cancer growth. • Geldanaymicn: initially thought to be due to specific tyrosine kinase inhibition, later studies revealed that the antitumor potential relies on the depletion of oncogenic protein kinases via the proteasome.
C. Hsp90 inhibitors cont. • Subsequent immunoprecipitation and X-ray crystallographic studies reveled that GA directly binds to Hsp90, and inhibits the formation of Hsp90 multichaperone complexes resulting in the ubiquitin-mediated degradation of Hsp90 client proteins. • This represented the first generation of drugs that specifically targeted Hsp90.
C. GA cont. • GA binds to the N-terminal domain of Hsp90 and competes with ATP binding. • The geldanamycin-Hsp90 crystal structure also shows that the binding inhibits substrate protein binding. • GA also binds to Grp94, the Hsp90 analogue in the ER
D. Advantages of Hsp90 inhibitors • Preclinical trials emphasize the important role of Hsp90 inhibitors in clinical applications. • Combination therapies, applying low doses of these drugs together with convention chemotherapeutic agents, seem to be an effective way to target various cancers. • For example, in the case of Bcr/Abl-expressing leukemias, a low dose GA is sufficient to sensitize these cells to apoptosis.
D. Advantages of Hsp90 Inhibitors • Among the hallmarks of cancer, up regulation of growth signals and evasion of apoptosis are the most important. • As most growth regulatory signals depend on Hsp90 for their function stability, Hsp90 is an ideal molecule to intervene in complex oncogenic pathways. • Hence, most drugs are targeting Hsp90, which is more beneficial than the selective oncogene pathway inhibitors.
E. 17-Allylamino, 17-Demethoxygeldanamycin (17-AAG) • An analog of GA. • There is a correlation between down-regulation of Hsp90’s client proteins and growth inhibition caused by 17-AAG. • 17-AAG has a higher affinity to bind to Hsp90 multichaperone complexes than Hsp90 in normal cells. There is only speculation of why this is true.
IV. Paper • “A high affinity conformation of Hsp90 confers tumor selectivity on Hsp90 inhibitors” • by: Kamal, Thao, Sensintaffar, Zhang, Boehm, Fritz, and Burrows
ImportantDiscoveries • “Tumour Hsp90 is present entirely in multichaperone complexes with high ATPase activity, whereas Hsp90 from normal tissue is in a latent uncomplexed state.” (Kamal 2003) • “In vitro reconstitution of chaperone complexes with Hsp90 resulted in increased binding affinity to 17-AAG, and increased ATPase activity.” (Kamal 2003)
Important Discoveries cont. • Tumour cells contain Hsp90 complexes in an activated, high-affinity conformation that promotes malignant progression, and that may represent a unique target for cancer therapeutics • How do we prove if Hsp90 from tumour cells had a higher binding affinity to 17-AAG than that from normal cells?
Figure 1: Results • “Hsp90 in tumour cells has a significantly higher binding affinity for 17-AAG than does Hsp90 from normal cells.” (Kamal 2003) • The binding affinity of 17-AAG to Hsp90 from the different cells directly correlates with the cytotoxic/cytostatic activity of 17-AAG in those cells (Fig. 1d)
Further Questions • Hsp90 interacts with many co-chaperone proteins that assemble in multi-chaperone complexes • Tumour cells overexpress Hsp90 client proteins • How to find out whether Hsp90 in tumour cells was present in multi-chaperone complexes?
Figure 2a: Results • “Co-immunoprecipitation in the normal and tumour cell lystates with antibodies to Hsp90 revealed that more Hsp90 in tumour cell was present in complexes with p23 and Hop compared to to normal cells” (Kamal 2003) • Control immunoprecipitaions without antibodies did not immunoprecipitate any Hsp90 • Immunoprecipitation with antibodies to both p23 and Hop revealed that the entire tumour cell pool of Hsp90 was present in complexes unlike normal cells.
Figure 2a: Results cont. • Immunoprecipitation with an antibody specific for the uncomplexed form of Hsp90 pulled down far more Hsp90 from normal cells than from tumour cells. • All Hsp90 in tumour cells is in the form multi-chaperone complex that are actively engaged in chaperoning client oncoproteins, and that the heightened complex formation is not due to increased expression of Hsp90 or co-chaperones.
Figure 2b: Results • Since the chaperone function of Hsp90 is dependent upon ATPase activity, they immunopreipitated Hsp90 from cell lysates and performed ATPase assays • “Tumour Hsp90 had markedly higher ATPase activity compared to Hsp90 from normal cells, and was inhibited by 17-AAG.” (Kamal 2003) • “Control immunoprecipitation in the absence of Hsp90 antibody did not have any significant ATPase activity (data not shown).” (Kamal 2003)
Figure 2: Combined Results • “These results suggest that essentially all soluble Hsp90 in tumour cells is present in fully active multi-chaperone complexes, whereas Hsp90 in normal cells is in an uncomplexed inactive form.” (Kamal 2003) • Data demonstrates that the Hsp90 from tumour cells had higher binding affinity to 17-AAG, and this correlates with presence of increased multi-chaperone complexes and increased ATPase activity. • Could there by any further tests?
Further Questions and Tests • How to examine whether reconstitution of purified Hsp90 with co-chaperones would result in increased binding affinity and ATPase activity? • In vitro reconstitution, they used five proteins that have been shown to be required for the in vitro chaperoning activity of Hsp90. (Hsp90, 70, 40, Hop, and p23)