
Картиране на генома - въведение
Картиране на генома - въведение. Една от основните задачи на проучването на генома е да се изследва секвенцията на геномните молекули.

Картиране на генома - въведение
E N D
Presentation Transcript
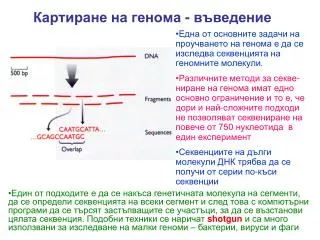
Картиране на генома - въведение Една от основните задачи на проучването на генома е да се изследва секвенцията на геномните молекули. Различните методи за секве-ниране на генома имат едно основно ограничение и то е, че дори и най-сложните подходи не позволяват секвениране на повече от 750 нуклеотида в един експеримент Секвенциите на дълги молекули ДНК трябва да се получи от серии по-къси секвенции • Един от подходите e да се накъса генетичната молекула на сегменти, да се определи секвенцията на всеки сегмент и след това с компютърни програми да се търсят застъпващите се участъци, за да се възстанови цялата секвенция. Подобни техники се наричат shotgun и са много използвани за изследване на малки геноми – бактерии, вируси и фаги
Картиране на генома - подходи • Техниката се усложнява с нарастване на генома. За n фрагмента възможният брой застъпвания 2n2 – 2n; • Друг проблем на техниката е при работа с повтарящи се секвенции в генома; • Когато се разрежат такива секвенции, много от фрагментите имат еднакви или сходни участъци (Фиг); • При това положение е много лесно да се възстанови погрешно цялата секвенция с липсващи участъци или такива, които не са на мястото си;
Картиране на генома – основни подходи • Разгледаните техники са неприложими при сложни геноми, като еукариотните. • Необходимо е да се състави геномна карта (генетична карта). • Тя показва местата на гените или на дискретни участъци на генома. Когато е има геномната карта, секвенирането може да протече по някой от следните два подхода: • Чрез clone contig (описан по-нататък). • Геномът се накъсва на фрагменти от няколкостотин килобази (kb) или на няколко мегабази (mb), които след това се секвенират отделно с shotgun метода. Когато се определи секвенцията на всеки сегмент, тя може да бъде позиционирана на даденото място в генома • Чрез директен shotgun подход (описан по-нататък). • Тази техника използва определени особености на геномната карта като маркери, за да се сглоби цялата секвенция от огромен брой къси секвенции, получени чрез shotgun техниката.
Маркери за генетичните карти • Както и при всички останали карти, една генетична карта трябва да показва положението на определени характерни особености, които по аналогия с географските карти се означават като маркери. • Основните маркери, които се използват за съставяне на генетични карти са гени или секвенции по веригата на ДНК
Гените като маркери – гени с фенотип • В началото на 20-ти век са конструирани първите генетични карти на дрозофилата, като генетични маркери са били използвани гените; • Гените са били разглеждани като абстрактни елементи, отговорни за пренасяне на наследствените белези от родител към поколение; • За да са полезни за съставяне на генетичната карта, тези белези трябва да съществуват в две форми – фенотипове, които се определят от алели на гена; • Такъв е например използваният от Мендел белег къси и дълги стъбла при граха. Всеки фенотип се характеризира от различен алел на съответния ген; • Има сравнително малко подобни визуални фенотипове, които биха могли да се използват за маркиране на генома, а освен това подобно маркиране не винаги е лесно, защото в някои случаи за определен белег отговарят повече от един ген; • Един от най-важните белези е бил цвета на очите, от които са различавани най-малко 10 нюанса на червения цвят (red, light red, vermilion, garnet, carnation, cinnabar, sepia, scarlet, pink, cardinalor claret), което определено означава, че цветът на очите на дрозофилата се определя от повече от един ген.
Маркери за генетичните картиГените като маркери на генетичните карти – гени без ясен фенотип • Очевидно е, че да се направи по-подробна генетична карта е било необходимо да се намерят по-голям брой белези, които да са по-ясно изразени и да са по-малко сложни, отколкото визуалните. • Отговорът на този въпрос е бил да се използват биохимични белези за отдиференцираненафенотиповете, нещо особено важно за два типа организми – микроорганизмите и човека. • При първите, каквито са бактерии, дрожди има много малко визуални характеристики, които биха могли да се използват за картиране, такаче генетичното им картиране очевидно трябва да се основава на био-химични фенотипове. • При хората съществуват визуални характеристики, но някои биохи-мичните фенотипове, като кръвногруповите характеристики са известни още през 20-те години на 20-ти век. • Изследвани са не само класическите АВО групи, но и алелните вариации на серумните белтъци и на някои имунни белтъци, каквито са човешките левкоцитни антигени, известни като HLA-система. • Едно съществено предимство на тези маркери пред визуалните фено-типове, е че много от свързаните с тях гени могат да са множествени алелни форми. Съществуването на множество алели в един локус се нарича генетиченполиморфизъм.
Маркери за генетичните картиНужда от други (ДНК) маркери на генетичните карти • Използването на гените е добър начин за картиране на генома, но когато се картират организми като гръбначни или висши растения той не е идеален • Основният проблем при подобни карти, е че те не са достатъчно детайлни. Дори да се картират всички гени, картата пак няма да е достатъчно подро-бна, защото тези гени са разделени от големи междугенни участъци. • Много гени имат алелни форми, които не могат да бъдат характеризирани с обикновени методи. • Дивият фенотип може да бъде полиморфен, без това да се отразява на фенотипа. Множествените форми на дивите алели могат да се разграничат чрез различия в тяхната секвенция, която не засяга тяхната функция, и по този начин не дава фенотипни варианти. В даден локус може да има много различни варианти на секвенциите – някои от тях са очевидни, защото за-сягат фенотипа, но други са скрити, защото нямат визуален ефект. • Следователно в един локус може да има множество промени, които:а) променят секвенцията на ДНК, но не променят секвенцията на белтъка;б) променят секвенцията на белтъка, без да променят функцията; в) дават белтъци с различаваща се активност; г) дават мутантни белтъци, които са неактивни. • Картиращите белези, които не са гени, се наричат ДНК маркери. Както и при гените, за да са използваеми, ДНК маркерите трябва да съществуват в поне две алелни форми. Има най-малко три типа секвенции ДНК, задоволява-щи тези изисквания, които ще бъдат разгледани по-нататък.
Маркери за генетичните карти – ДНК маркериПолиморфизъм в дължината на рестрикционни фрагменти (Restriction Fragments Length Polymorphism - RFLPs) Молекулата ДНК отляво има полиморфен рестрикционенсайт (*), който липсва в молекулата отдясно. RFLP се установява след третиране с рестриктазен ензим, защото едната ДНК се реже на четири, а другата на три фрагмента. • RFLPs са първите използвани ДНК маркери. Те се получават с рестрикционни ензими, които режат молекулата на ДНК на места с определени секвенции • Някои от рестрикционните сайтове са полиморфни, съществуващи в два алела, така че ако единият алел има рестрикционния сайт, а другият – не, той не се разрязва от ензима • Резултатът е, че двата съседни рестрикционни фрагмента ще останат свързани заедно след разрязването с ензима, което ще доведе до полиморфизъм в дължината. • Това се нарича RFLP и неговото място по генетичната карта се установявас унасле-дяването на алелите, точно както когато гените се използват като генетични маркери. • Полиморфизъм на това ниво може да се установи независимо дали засяга фенотипа. Вероятно много малко от полиморфните рестрикционни сайтове засягат фенотипа.
Маркери за генетичните карти – ДНК маркериПолиморфизъм в дължината на прости секвенции(Single Sequences Length Polymorphism - SSLPs) Типизиране на SSLPs. (А) Два алела на микросателита SSLP. В алела 1 мотивът GA се повтаря три пъти, а в алела 2 – пет пъти. (В) Как SSLP може да се типизира чрез PCR ? Областта, заобикаляща SSLP се амплифицира и продуктът се изследва чрез агарозна електрофореза. Продуктите на PCR дават зона, отговаряща по големина на най-голямата от двете алелни сек-венции, което показва, че изследваната ДНК съдържа алел 2. Съществуват два типа SSLP: Минисателити, известни също като променлив брой тандемни повтори (Variable Number of Tandem Repeats -VNTRs), в които повтарящата се секвенция е дълга няколко десетки нуклеотида (GAGAGAGAGA) Микросателити или прости тандемни повтори (Simple Tandem Repeats – STRs), чиито повторения са значително по-къси, обикновено ди-, три- или тетрануклеотидни единици (GAGA).
Маркери за генетичните карти – ДНК маркериПолиморфизъм на единични нуклеотиди(Single Nucleotide Polymorphism – SNPs) • Съществуват много точкови мутации в генома, някои от които дават RFLPs, но много други нямат такъв резултат, защото не са в места, които се разпознават от рестриктази. Счита се, че в човешкия геном има 200 000 такива SNPsв отделните гени и може би десет пъти повече в негеномната ДНК. • Всеки SNP може да е само в две алелни състояния, което означава, че този метод има същите недостатъци, които и RFLPs тъй като има висока вероятност за хомозиготност на човешките семейства по отношение на дадени SNPs. • Предимствата на метода са, че SNPs са много голям брой и освен това те могат да се откриват с методи, които не изискват електрофореза. Това е важно, защото този метод не се подава на автоматизиране и всички техники, които използват електрофореза ще са относително бавни. Откриването на SNPs е по-бързо, защото се базира на олигонуклеотиден хибридизационен анализ, която се означава още като технология на ДНК чип (microarray).
SNPs се откриват чрез: • Технология на ДНК чип. Това е силициева пластина с размери 2Х2 cm, върху която са подредени с висока плътност различни олигонуклеотиди. Пластината се третира с тестовия олигонуклеотид, който е белязан с флуоресцентен маркер, а хибридизацията се открива с флуоресцентен микроскоп, който сканира повърхността на пластината. Това означава тестване на голям брой ДНК за кратко време. • Динамична алелно-специфична хибридизация.Хибридизацията се извършва в разтвор, и се открива по флуоресцентен маркер, който се свързва само с двойноверижна ДНК. Хибридизацията се извършва при условия, при които хибридизират всички молекули, дори тези, които имат един различен нуклеотид. Дискриминацията става по-късно, когато се повишава температурата и тогава двуверижен остава само “идеалният” хибрид
Подходи на генетичното картиране • При мейозата двойките хомоложни хромозоми не са самостоятелни. • По време на мейоза I всяка хромозома образува с хомолога си бивалент. • Това става след като всяка хромозома се е реплицирала, но преди реплицираните структури да се отделят, така че бивалента носи четири копия от хромозомата, всяко от които ще отиде в една от гаметите. • В бивалента хромозомните рамене (хроматидите) могат да претърпят физични прекъсвания и обмени на участъци от ДНК. • Този процес е известен като кросинговър или рекомбинация и е открит от Янсенс през 1902 г., само две години преди Морган да започне да мислиза частичната скаченост.
Алтернативи при кросинговъра • Съществуват две алтернативи: • Между гените А и В няма кросинговър. Тогава получените гамети ще съдържат хромозомни копия с алелите А и В, а останалите две гамети – с a и b. С други думи две от гаметите ще имат генотип АВ, другите де – ab. • Между гените А и В се наблюдава кросинговър. Това ще доведе до поява на сегменти ДНК, съдържащи гена В, обменен между хомоложните хромозоми. Това ще доведе до поява на гамети с различен генотип: 1 AB, 1 aB, 1 Ab, 1 ab.
Анализ за скаченост (linkage analyses) - основа на генетичното картиране • Основните техники за картиране се основават на анализ за скаченост (linkage analyses), открит през 1905 г. от Бейтсън, Сандерс и Панет. Той остава неясен, докато Морган не започва да работи с дрозофилата през 1910-11. • Тогава се счита, че ако два гена са разположени в една хромозома, то те са физически свързани (скачени) и затова трябва да се унаследяват заедно • Мендел установява, че пълна скаченост показват само малка част от гените. Една част от двойките гени се унаследяват самостоятелно, както би могло да се очаква ако са гени от различни хромозоми, което означава, че те проявяват частична скаченост, тъй като понякога те се унаследяват заедно, понякога отделно. • Разрешаването на това несъответствие между теория и практика всъщност поставя основата на техниките на генетичното картиране
Частична скаченост и генетично картиране След обясняване на механизма на частичната скаченост чрез кросинговър, Морган успява да създаде начин за картиране на относителното положение на гените в хромозомата. Всъщност пробива в това направление е направен от един студент - Артър Стъртивент (Sturtevant, 1913), който предполага, че кросинговърът е случайно събитие и той може да стане с равна вероятност между всеки участък на двата хроматида. Ако това е вярно, то два гена, които са близо един до друг, ще бъдат разделяни от кросинговъра по-рядко от тези, които са на по-голямо разстояние. Така честотата, с която гените са “разкачвани” един от друг ще е право пропорционално на разстоянието, на което се намират в хромозомата. Това се нарича честота на рекомбинация и всъщност е мярка за разстоянието между гените. Ако се измери честотата на рекомбинация за различни двойки гени, може да се конструира карта на относителното им положение върху хромозомата. Генетична карта, съставяна чрез честота на рекомбинация. На фигурата е показан оригиналният експеримент на Sturtevant. Всичките четири гена са на X хромозомата на дрозофилата. Показана е честотата на рекомбинациямежду гените и предполагаемото им картиране.
Анализ на скаченост при организми, при които са възможни планирани експерименти на кръстосване Два примера на тестово кръстосване: По сценарий 1, А и В са генетични маркери с алели A, aи B, b. Полученото потомство се изследва на базата на техния фенотип. Тъй като двойния хомозиготен родител (Parent 2) има и двата рецесивни алела (a и b) той не влияе на фенотипа на поколението. Така фенотипа на всеки индивид от F1 е еднакъв с генотипа на гаметите от другия родител (Parent 1), които са го създали. По сценарий 2, А и В са ДНК маркери, чиито алелни двойки са кодоминантни. В конкретния случай, двойният хомозиготен родител (Parent 2) има генотипа Ab/Ab. Алелите, които присъстват във всеки индивид се тестват пряко, например чрез PCR. Това алелно кодоминиране дава възможност да се предположи генотипа на гаметата на първия родител (Parent 1), който дава всеки индивид
Картиране на гените чрез изследване на родословни дървета при човека • При човека не могат да се подбират генотипове на родителите и да се правят планирани кръстоски за картирането. • Честотата на рекомбинация трябва да се изследва с генотиповете на членовете на поколения от съществуващи семейства. • Най-често има само ограничени данни и тяхното интерпретиране е трудно, тъй като човешките семейства не приличат на тестово кръстосване. • Генотиповете на някои от членовете на семей-ството могат да липсват - починали или пък не желаят да сътрудничат в такива експерименти.
Картиране на гените чрез изследване на родословни дървета при човека • (А) Унаследяването на генетично заболяване от два живи родителя и 6 деца с информация за прародителите по майчина линия. Болният алел е доминантен над здравия. Да се определи степента на скаченост на болния ген и микросателита М чрез типизиране на алелите на този микросателит (М1, М2 и т.н.) в живите членове на семейството. • (В) Родословното дърво може да се интерпретира по 2 начина. • Ниска честота на рекомбинация и ген на заболяването, тясно свързан с микросателита М; • Гена и микросателита са в много по-малка степен скачени. • (С) Случаят може да се разреши с появата на бабата, чийто генотип микросателити отговаря на хипотеза1.
Картиране на гените чрез изследване на родословни дървета при човека • Генетичните заболявания се използват често като генни маркери при човека, като болестта е един алел, а липсата на заболяване – друг алел. • Схемата показва, че майката е засегната от заболяването, както и 4 от нейните деца. • Известно, че бабата на майката също е страдала от това заболяване, но тя и нейният съпруг са починали, без да има данни за геномите им. • Целта е да се картира гена, причиняващ генетичното заболяване и се изследва неговата скаченост с генетичния маркер М, като в живите членове на семейството има четири негови алела – M1, M2,M3 и M4. • Въпросът е колко от децата в семейството са рекомбинанти?
Картиране на гените чрез изследване на родословни дървета при човека • Вижда се, че децата 1, 2 и 4 имат болестния алел и микросателитния алел M1. • Децата 2 и 5 имат здрав алел и M2. Можем да конструираме две алтернативни хипотези: • 1. Двете копия на съответната двойка хомоложни хромозоми у майката имат генотипове болен-M1, здрав-M2 и затова децата 1, 2, 3, 4 и 5 имат родителските генотипове и само шестото дете е рекомбинант (В). Това може да предполага, че болния ген и микросателита са скачени сравнително близо и рекомбинацията между тях става сравнително рядко. • 2. Майчините хромозоми имат генотип болен-M2, здрав-M1, а децата 1-5 са рекомбинанти, а шестото дете има родителския генотип. • Това означава, че гена и маркера са относително далеч един от друг в хромозомата и кросинговър става по-често. • Данните от това семейство не са достатъчни, за да преценим коя от двете хипотези е вярната.
Картиране на гените чрез изследване на родословни дървета при човека • Разрешението на проблема: • Ако знаем генотипа на бабата (жива) и че генотипа за микросателит М е М1М5. • Това показва, че болестния алел се намира на същата хромозома, в която М1. • Така със сигурност можем да кажем, че хипотеза 1 е вярната и само шестото дете е рекомбинант.
Генетично картиране при бактериите (1) • Основната трудност при картирането на бактериите е, че са хаплоидни организми и при тях няма мейоза. Трябва да се търсят други начини за кросинговър между хомоложни сегменти на бактериалната ДНК. Такива са трите известни начина, по които става прехвърляне на генетичен материал от една бактерия в друга: • Конюгация - двете бактерии са във физически контакт, и едната бактерия (донор) предава генетична информация на другата (реципиент). Това става през тясна тръба (пилус). Прехвърлената ДНК може да е част или дори всички бактериални хромозоми, или фрагмент от бактериалната ДНК с дължина до 1 Mb, интегриран в плазмид. Последният начин се нарича още епизомен трансфер и се извършва при репликация на хромозомата на донорната бактерия по търкалящия се пръстен. • Трансдукция, която представлява прехвърляне на малки сегменти до 50 kb от бактериалната ДНК от донора в реципиента чрез бактериофаги. • Трансформация, при която клетките приемат от външната среда сравнително малки фрагменти ДНК (рядко, повече от 50 kb), отделени от донорните клетки
Генетично картиране при бактериите (2) • След прехвърлянето трябва да се извърши двоен кросинговър, за да може ДНК от донора да се интегрира в хромозомата на реципиента. Единственото изключение е епизомния трансфер, тъй като плазмидите могат да се делят независимо от бактериалната хромозома. • Обикновено за анализа се използват биохимични маркери, като доминантния или див тип на фенотипа е биохимична характеристика (може да синтезира триптофан или чувствителност към антибиотик), а рецесивния фенотип елипсата им. • Преноса се прави между донорен щам с алели от дивия тип и рецепиентен щам с рецесивни алели, а прехвърлянето се следи по появата в реципиента на дивия фенотип. • Процедурата зависи от начина, по който се прехвърля на ДНК. Например при конюгацията се следи времето, за което дивия алел на гена навлиза в реципиента. • Трансдукцията и трансформацията позволяват картиране на близки гени, тъй като прехвърляните ДНК са относително къси. • Вероятността два гена да се прехвърлят заедно зависи от разстоянието между тях на бактериалната хромозома.
Физично картиране на генома • Генетичният анализ може да се използва за съставяне на генетични карти за локализация на гени или генетични маркери като RFLP, SSLP и SNP върху хромозомите. Една алтернатива на генетичното картиране е прякото физично изследване на генома, основаващо се на директно изследване на молекулата ДНК. • Интересно е да се сравнят двата типа карти – генетичните и физичните и степента на тяхното съвпадение. Те биха могли да се допълват и така да се получи една цялостна картина за генома. Нуждата от физично картиране на генома е породена от някои недостатъци на генетичните карти, които биха могли да се обобщят в две основи насоки – ниска разделителна способност и малка точност. • Генетичните карти имат ограничена разделителнаспособност: • Микроорганизмите могат да се отглеждат в много големи количества и да се получават много рекомбинанти. Това дава възможност за картиране на много близки участъци разделени само с няколко килобази. Генетичната карта на E. coli е съставена от 1 400 маркераи средно разстояние между тях 3,3 kb - достатъчно за секвениране, без да се налага физично картиране. • Проблемът е при картиране на човешкия геном, където не могат да се изследват много поколения. Днешната разделителна способност на генетичното картиране 0.6 Mb, е недостатъчна за човешкия геном • Генетичните карти имат ограничена точност. Рекомбинациите стават на случайни места по хромозомите, но някъде те стават по-често. Ако се сравни генетичната карта със секвенирания геном се вижда, че една двойка гени дори са разменени от генетичния анализ. Като се има предвид, че генома на дрождите е един от най изучените и ако генетичната карта на дрождите е неточна, то какво остава за останалите, по-малко проучени, включително и на човека.
За физичното картиране са разработени поредица от методи, които могат да се разделят на три основни групи: • Рестрикционнокартиране, с което върху молекулата на ДНК се определя относителното положение на местата на рязане на рестрикционни ензими. • Флуоресцентна хибридизация in situ(FISH), при която локализацията на маркерите се картира чрез хибридизацията им върху интактната хромозома. • Картиране чрез маркиране на секвенции на сайтове (STS – sequence tagged site mapping), позицията на къси секвенции се маркира чрез PCR и/или хибридизационен анализ на геномни фрагменти.
Рестрикционно картиране Рестрикционни ензими: • Сайт-специфични ендонуклеази • Дават “лепливи” и “тъпи” краища • Характерни са за всяка бактерия • Имат различни по дължина на нуклеотидите “таргетни”сайтове на рязане • Дават различни по дължина фрагменти • Малък брой от тях притежават генетичен полиморфизъм Основни методи на рестрикционно картиране: • Сравняване на дължината на получените фрагменти при рязане на една ДНК с два различни ензима (напр. EcoR1 и BamH1) – двойно рязане • Частично рязане на ДНК с рестрикционни ензими – дава края на рестрикционната карта
Рестрикционното картиране – ограничено от дължината на получените рестрикционни фрагменти • Рестрикционните карти се правят сравнително лесно, когато в ДНК има малко рестрикционни сайтове на ензима. • С нарастването на местата на рязане, нараства и броят на единичните, двойните и частично разрязани фрагменти, които трябва да се сравнят и определят, за да се състави генетичната карта. • Въпреки използваният компютърен анализ, се стига до етап, когато фрагментите се сливат, увеличавайки шанса все повече фрагменти да бъдат измерени неточно или въобще да се пропуснат. • Но дори и всички да бъдат идентифицирани, възможно е няколко да имат сходни размери и това да не позволи разполагането им на една непрекъсната рестрикционна карта.
Рестрикционното картиране – ограничено от дължината на получените рестрикционни фрагменти - продължение • Рестрикционното картиране е приложимо повече за сравнително къси ДНК, като горната граница зависи от използваните рестриктази. • Ако ДНК е по-къса от 50 kb, съставянето на рестрикционна карта не е проблем с използване на ензими, разпознаващи 6-нуклеотидни сайтове. • Това е по-малко от генома на бактериите и близо до този на фагови и вирусни геноми, както и на геномите на органелите в клетката. • Рестрикционните карти са полезни, когато бактериални или еукариотни геноми са клонирани, тъй като клоновете не са по-дълги от 50 kb и може да се направи подробна карта на клона, преди да бъде секвениран.
Рестрикционни карти на геноми, по-големи от 50 kb: Ензими, които режат ДНК на големи разстояния (rare cutters) • Някои рестрикционни ензими, които разпознават дълги секвенции от седем или осем нуклеотида. Това са сравнително малък брой ензими. • Такива са например SapI (5΄-GCTGTTC-3΄) SgfI (5΄-GCGATCGC-3΄). • Седем-нуклеотидните ензими би трябвало да режат ДНК с 50% съдържание на GC с честота около 47 = 16 384 bp, а осем-нуклеотидната рестриктаза – 48 = 65 536 bp. • За сравнение шест-нуклеотидните ензими, каквито са BamH1 иEcoR1 дават фрагменти 46 = 4096 bp. • По-рядко режещите ензими се използват за кариране на големи геномни ДНК но подходът не е много използван, защото подобни ензими са рядкост.
Ензими, разпознаващи секвенции, които са редки в дадена ДНК (rare cutters) • Геномните ДНК рядко имат съвсем случайни секвенции, като някои от тях не притежават определени групи (мотиви). • Така секвенцията 5΄-CG-3΄е рядка за човешката ДНК, тъй като в клетките има ензим, който метилира 5΄-въглерода на цитозина. • Това дава 5-метилцитозин, който има тенденция да се деаминира и да се превръща в тимин. В резултатът по времена еволюцията много от мотивите 5΄-CG-3΄ са се превърнали в секвенции 5΄-ТG-3΄. • Затова рестриктазите, които имат разпознаващ сайт с 5΄-CG-3΄, режат човешката ДНК по-рядко. • Такъв е ензима SmaI (5΄-CCCGGG-3΄),който реже човешката ДНК на всеки 78 kb, както и BssHII (5΄-GCGCGC-3΄), който режена всеки 390 kb. • Някои rare cutters, като NotI, също имат5΄-CG-3΄ в разпознаващия сайт и режат човешка-та ДНК много рядко – на всеки 10 Mb.
Подходи за определяне на големи ДНК, с ензими rare cutters – обща характеристика • При използването на тези ензими се получават големи фрагменти ДНК, които се разделят трудно с конвенционална електрофореза в агароза. • След определена дължина (над 50 kb) фрагментите започват да се движат с еднаква скорост и образуват бавно придвижваща се зона. • За да се разделят големи фрагменти се използват модификации на електрофорезата • Такъв метод е прилагането на променящо се електрично поле между две двойки електроди, разположени под ъгъл 45о на гела, което кара по-малките фрагменти да се движат по-бързо.
Подходи за определяне на дългите фрагменти ДНК, с ензими rare cutters – оптично картиране Отчитането на получените фрагменти може да става и с други техники. • Изолирани хромозоми от дрожди са оптично картирани, при което рязането на ДНК се наблюдава с мощен флуоресцентен микроскоп. • Необходимо е да се получат изпънати нишки ДНК, които да се подложат на действието на рестриктазата. • Методът се нарича разпъване на гела. Високомолекулната ДНК се смесва с агарозен гел, той се излива на предметно стъкло и при застиване и втвърдяване му молекулите ДНК се изпъват, защото слоят е много тънък. • Рестриктазата се нанася върху стъклото, а ДНК се оцветява с DAPI (4,6-диамино-2-фенилиндол дихидрохлорид). • Бавно между разрязаните фрагменти се получават разстояния поради свиването на спиралите на ДНК и под микроскопа се регистрират местата на рязане и дължината на фрагментите.
FISH – флуоресцентна хибридизация in situ • Свързано е донякъде с оптичното картиране (ОК), като FISH позволява оптично картиране на местата на маркерите по хромозомата. • Вместо визуализиране на рестрикционни сайтове (ОК), при FISH маркерите са определени секвенции ДНК и наблюдението им става след хибридизация с флуоресцентна сонда. • Хибридизация in situ с използване на радиоактивни или флуоресцентни проби • Хибридизацията in situ е метод, при който хромозомата се изследва чрез хибридизация с белязани молекули ДНК. • Мястото на хибридизацията върху хромозомата картира мястотона секвенцията, използвана като проба. • За целта ДНК на хромозомата трябва да е едноверижна, което става с денатурация на ДНК и едва тогава се извършва хибридизация. • Денатурация на ДНК става при стайна температура като тя се изсушава върху предметно стъкло и препаратът се обработи с формамид (намалява се Тm).
FISH – флуоресцентна хибридизацияin situ - продължение • При отчитането на хибридизация се използват радиоактивни проби, като зацелта има методи за радиоактивно белязане на ДНК. • Може да се използва и нерадиоактивното флуоресцентно белязане, при което се комбинира високата чувствителност с висока разделителна способност и са идеални за in situ хибридизация. • При нерадиоактивното белязана се използва цветна, чувствителна биохимична реакция, а не отчитане на радиоактивност. • Нерадиоактивното белязане е по-малко чувствително от радиоактивното.
In situ хибридизация, продължение • При ползването на дълги фрагменти ДНК като проби има проблем, свързан с повтарящи се секвенции в пробата. • Пробите могат да се свързват не само на едно място по хромозомата със сходство на секвенциите, но и на места с по-слаба хибридизация, но все пак достатъчна за формиране на дуплекси. • За намаляване на тази неспецифична хибридизация, първоначално пробата се смесва с небелязаната ДНК, която ще бъде изследвана. • Идеята е, че небелязаната ДНК хибридизира с повтарящите се участъци в пробата и ги блокира, така че последвалата in situ хибридизация се осъществява само от уникалните секвенции и неспецифичната хибридизация се елиминира.
Същност на хибридизацията in situ FISH с ниска разделителна способност при метафазни хромозоми • Първоначално FISH се използва с метафазни хромозоми, при които флуоресцентният сигнал се картира чрез измерване разстоянието спрямо късото рамо на хромозомата, известно като стойност FL(FLpter value). • Метафазните хромозоми са силно кондензирании така може да се направи карта с ниска разделителна способност. • Приложението на метафазнатаFISH е било да се определи на коя хромозома е маркера или грубо да се прецени неговото картиране, което може да е основа на по-фини карти. FISH с висока разделителна способност с дълги хромозоми и ДНК. • Резултатът от хибридизацията in situ се определя основно от подготовката на хромозомата, като основният проблем е високата степен на организация на ДНК в метафазните хромозоми. • Успехът на метода е в намаляване на компактизацията на ДНК и на самата хромозома. Това се постига по два основни начина:
Механично изпъване на хромозомите. • Може да се постигне с центрофугиране, като хромозомата се изпъва около 20 пъти. • При това хромозомите все още могат да се различават, а FISH сигналът може да се картира по същия начин, както и при метафазните хромозоми. • Разделителната способност нараства значително и така могат да се картират секвенции само на 200-300 kb една от друга.
Неметафазнихромозоми • Използвани са профазни хромозоми, които са вече достатъчно кондензирани, за да се различават. • Методът няма особени предимства пред механичното изпъване. • Използвани са и интерфазните хромозоми, които са най-малко пакетирани. • Достигната е разрешителна способност до 25 kb, но тези хромозомите са неразличими, което пречи за картирането. • Методът се използва след предварителна информация за картата, която се нуждае от детайлизиране и за откриване на серии от маркери на малки участъци от хромозомата.
Други подходи • Най-висока разделителната способност на FISH се постигна чрез използване на пречистена ДНК, която се изпъва в гел (fiber-FISH). • Друг подход е молекулен “гребен” от нишки ДНК, който се прави чрез потапяне на стъкло, покрито със силиций, в разтвор на ДНК. • При изваждането му силата, необходима да издърпа нишките ДНК през менискуса на повърхността на разтвора ги подрежда успоредно една на друга (“гребен”) и като изсъхнат на въздуха, те са успоредно подредени молекули ДНК. • Така се картират маркери на разстояние около 10 kb.
Нужда от нови подходипри физичното картиране • Както стана ясно основните изисквания на физичното картиране на генома са бързина и ниска техническа сложност. • И двата споменати досега метода не отговарят на тези изисквания. Какви са недостатъците им? • Рестрикционното картиране е лесно за изпълнение и има достатъчна чувствителност, но с него могат да се картират само малки геноми. • Използването на FISH позволява картиране на големи геноми, a модификации като fiber-FISH позволяват сравнително висока разделителна способност, но е трудна за изпълнение и позволява максимално картиране на 3-4 маркера за един експеримент. • Ако трябва да се направят детайли физични карти на генома са нужни значително по-съвършени методи.
Картиране със сайтове, маркирани по дадена секвенция (sequence tagged sites – STS) • Днес най-усъвършенстваният метод за физично картиране на големи геноми е STS. • Маркираният по секвенция сайт, или STS, представлява къса секвенция ДНК, обикновено от 100 до 500 bp, която е лесна за разпознаване (секвениране; хибридизация) и се среща само веднъж в хромозомата или изследвания геном. • За да се картира група STS е необходимо от хромозомата или целия геном да се създаде серия от припокриващи се фрагменти ДНК. • Използва се колекция фрагменти от една хромозома, върху която има два близко разположени и два далечно разположени маркери. • Анализът на тяхното разположение се прави, като се изследват отделните фрагменти и се отбелязва кой фрагмент какъв маркер съдържа.
Картиране със сайтове, маркирани по определена секвенция (STS) (2) • Това може да се направи с хибридизация, но най-често в последно време се използва PCR, тъй като става по-бързо и се подава на автоматизация. • В зависимост от разстоянието между маркерите в генома, тези, които са близко разположени имат по-голям шанс да попаднат заедно на един фрагмент; • Ако са много близо, те винаги ще са заедно, а ако са по-далеч, понякога ще бъдат на един фрагмент, понякога не. • Данните могат да се използват, за да се изчисли разстоянието между тях както това се прави с анализ на скаченост, когато се изчислява честотата, с която между двата маркера се наблюдава кросинговър. • STS анализът е аналогичен, само че тук се изчислява честотата, с която между маркерите се наблюдават разкъсвания.
STS картиране • Колекция от фрагменти, подходяща за STS картиране. Фрагментите се простират по цялата дължина на хромозомата, като всяка точка на хромозомата е представена средно в 5 фрагмента. Двата маркера са близо на хромозомната карта и има голяма вероятност те да се намерят в един фрагмент. Други два маркера са по-далеч един от друг и затова е по-малко вероятно да се открият на един фрагмент.
Природа на STS • За да изпълни изискванията на STS, секвенцията ДНК трябва да отговаря на две основни изисквания: • Първо - секвенцията да е известна, което да позволява да се използва PCR за тестване дали секвенцията присъства на дадена ДНК. • Второ – STS трябва да са локализирани на уникални места по изследваната хромозома или на генома като цяло, ако подготвяния комплект фрагменти е от целия геном. • Ако секвенцията STS се наблюдава на няколко места, тогава картирането на обекта става нееднозначно. • Това означава, че STS не могат да бъдат в участъците с повтаряща се ДНК. • Тези критерии могат да бъдат задоволени сравнително лесно и STS могат да се получат по няколко начина:
Белези, от експресиращи се секвенции. Еxpressed Sequences Tags, EST • Това са къси секвенции, получени от анализа на комплементарна ДНК или копиДНК (cDNA), която се получава чрез копиране на иРНК в двойноверижна ДНК, което е достатъчно рутинна операция в молекулярната генетика (фигура). • Тъй като иРНК в клетката се получават от белтък-синтезиращите гени, това означава, че това са експресиращи се гени (ДНК). • В този случай ЕST се разглеждат като бърз начин за получаване на достъп до секвенции от важни гени, при което те са значими дори тяхната секвенция да не е пълна. • Една такава EST може да се използва като STS, когато е получена от уникален ген, който не принадлежи на семейство гени, в което гените имат много близка секвенция.
Метод за приготвяне на сДНК с използване на поли-А опашката на еукариотните иРНК • Първия етап е свързан с образуване на праймер върху поли-А от обратна транскриптаза, който продължава с копиране на цялата верига. • Следва разрушаване на веригата на иРНК с рибонуклеаза Н. • Втората ДНК верига се синтезира от ДНK полимераза I.
Други STS • SSLPs. • Това са секвенции за генетично картиране, които бяха разгледани по-рано, които биха могли да е използват също и като STS във физичното картиране. • SSLPs, които са полиморфни и вече са картирани чрез анализ на скаченост са особено полезни, тъй като те осигуряват връзка между генетичните и физични карти на генома. • Случайни геномни секвенции. • Получават се чрез секвениране на случайни участъци от клонираната геномна ДНК, или просто чрез търсене на секвенции, представени в различни бази от данни.
Получаване на фрагменти от ДНК за картиране с STS. Радиациационни хибриди • Това са клетки от мишки, които съдържат фрагменти от хромозоми от друг организъм. • Техниката е разработена още в 70-те години на 20-ти век, когато става ясно, че при излагане на човешки клетки на рентгенови лъчи с дози от порядъка на 3000-8000 рада предизвиква разкъсване на хромозомите на случайни фрагменти, като по-големи дози дават по-малки фрагменти. • Тези дози са смъртоносни за клетките, но ако те бъдат слети с други клетки (хамстер), като фузията става с химично въздействие, полиетиленгликол или под въздействие на вируса на Сендай, хибридните клетки могат да задържат определени фрагменти от човешките хромозоми. • Това не става винаги, затова е необходим метод за проверка на хибридите.
Mетод за проверка на хибридите • Такъв рутинен селекционен процес е използването на клетки от хамстер, които не могат да синтезират тимидин киназа (ТК) или хипоксантин фосфорибозил трансфераза (HPRT). • Липсата на тези два ензима е летална, когато клетките се отглеждат в среда, съдържаща смес от хипоксантин+птеридин+тимидин (НАТ-среда). • Клетките, които оживяват на НАТ-среда са хибридни, тъй като те са получили нормални ТК и HPRT гени от човешките хромозомни фрагменти, които им позволяват да разграждат НАТ-средата. • Получените хибридни клетки представляват популация, съдържаща случайни фрагменти от човешки хромозоми, които са инсертирани в хромозомите на хамстера. • Обикновено това са фрагменти от около 5-10 Mb, като всяка клетка съдържа около 15-35% от човешкия геном. • Колекцията от такива клетки се нарича панел от радиационни хибриди и те могат да се използват като реактивза STS картирането, ако се вземат мерки PCR реакцията, която се използва да не намножи фрагменти от хромозомите на хамстера.
Други панели от хибриди • Може да се конструира и друг тип панел от хибриди, съдържащи ДНК само от една човешка хромозома, като клетката, която се облъчва не е човешка клетка, а хибридна клетка, получена, както е описано по-горе. • Получени са голям брой клетки от мишки, в които нормално се реплицира само една човешка хромозома. • Ако такава клетка се облъчи с рентгенови лъчи и хибридизира с клетки от хамстер, то хибридите ще съдържат фрагменти от хромозоми от човек или мишка или пък смес от двете. • Тези, които съдържат човешка ДНК могат да се идентифицират чрез специфични сонди, като например къси, повтарящи се секвенции (SINE, short interspersed nuclear elements) - Alu-секвенции. • За тях е известно, че се повтарят около един милион пъти, което означава, че в човешкия геном те се появяват веднъж на всеки 4 kb. • Само клетки, които съдържат човешка ДНК ще хибридизират с Alu-секвенциите, което позволява да се отстранят мишите хибриди и STS секвенирането да се насочи към клетките, които съдържат човешки хромозомни фрагменти.





















