Download
1 / 12
150 likes | 603 Views
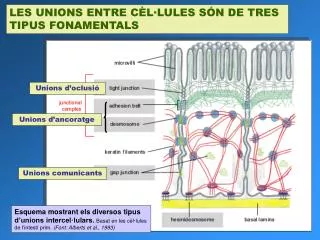
LES UNIONS ENTRE CÈL·LULES SÓN DE TRES TIPUS FONAMENTALS. Unions d’oclusió. Unions d’ancoratge. Unions comunicants. Esquema mostrant els diversos tipus d’unions intercel·lulars. Basat en les cèl·lules de l’intestí prim. (Font: Alberts et al., 1993).

E N D
LES UNIONS ENTRE CÈL·LULES SÓN DE TRES TIPUS FONAMENTALS Unions d’oclusió Unions d’ancoratge Unions comunicants Esquema mostrant els diversos tipus d’unions intercel·lulars.Basat en les cèl·lules de l’intestí prim. (Font: Alberts et al., 1993)
Principals tipus de molècules d’adhesió intercel·lular Interaccions homofíliques Interaccions heterofíliques Figure 22-2. Major families of cell-adhesion molecules (CAMs). Integral membrane proteins are built of multiple domains. Cadherin and the immunoglobulin (Ig) superfamily of CAMs mediate homophilic cell-cell adhesion. For cadherin, calcium binding to sites (orange) between the five domains in the extracellular segment is necessary for cell adhesion; the N-terminal domain (blue) causes cadherin to dimerize and to bind cadherin dimers from the opposite membrane. The Ig superfamily contains multiple domains (green) similar in structure to immunoglobulins and frequently contain type III fibronectin repeats (purple). In a heterophilic interaction, the lectin domain of selectins binds carbohydrate chains in mucin-like CAMs on adjacent cells in the presence of Ca2+. The lectin domain is separated from the membrane by a series of repeated domains. The major cell-matrix adhesion molecule, integrin, is a heterodimer of α and β subunits. They bind to the cell-binding domain of fibronectin, laminin, or other matrix molecules. (Font: Lodish et al., 2000) Figure 19-26. Three mechanisms by which cell-surface molecules can mediate cell-cell adhesion. Although all of these mechanisms can operate in animals, the one that depends on an extracellular linker molecule seems to be least common. (Font: Alberts et al., 1993)
Les unions d’ancoratge connecten el citoesquelet de les cèl·lules veïnes Hi ha dos tipus d’ unions d’ancoratge: unions adherents i desmosomes Les unions adherents connecten feixos de fibres d’actina entre cèl·lules Figure 22-5. Adhesion molecules in junctions involved in cell-cell adhesion. Adherens junctions and desmosomes are specialized cell-cell junctions that consist of clustered-cadherin dimers. Cadherin is connected to either the circumferential belt of actin filaments or bundles of keratin filaments in the cytoskeleton through the catenin adapter proteins. (Font: Lodish et al., 2000) Figure 19-8. Adhesion belts between epithelial cells in the small intestine. This beltlike anchoring junction encircles each of the interacting cells. Its most obvious feature is a contractile bundle of actin filaments running along the cytoplasmic surface of the junctional plasma membrane. The actin filaments are joined from cell to cell by transmembrane linker proteins (cadherins), whose extracellular domain binds to the extracellular domain of an identical cadherin molecule on the adjacent cell.(Font: Alberts et al., 1993)
Els desmosomes connecten els filaments intermedis de les cèl·lules veïnes Figure 22-6. Desmosomes. (a) Schematic model showing components of a desmosome between epithelial cells and attachments to the sides of keratin intermediate filaments, which crisscross the interior of cells. The transmembrane linker proteins, desmoglein and desmocollin, belong to the cadherin family. (b) Electron micrograph of a thin section of a desmosome connecting two cultured differentiated human keratinocytes. Bundles of intermediate filaments radiate from the two darkly staining cytoplasmic plaques that line the inner surface of the adjacent plasma membranes. [Part (a) see B. M. Gumbiner, 1993, Neuron11:551; D. R. Garrod, 1993, Curr. Opin. Cell Biol.5:30] (Font: Lodish et al., 2000)
Tant unions adherents com desmosomes utilitzen el mateix tipus de proteïnes per l’adhesió intercel·lular: les cadherines Figure 19-24. Schematic drawing of a typical cadherin molecule. The extracellular part of the protein is folded into five similar domains, three of which contain Ca2+-binding sites. The extracellular domain farthest from the membrane is thought to mediate cell-cell adhesion; the sequence His-Ala-Val in this domain seems to be involved, as peptides with this sequence inhibit cadherin-mediated adhesion. The cytoplasmic tail interacts with the actin cytoskeleton via a number of intracellular attachment proteins, including three catenin proteins. α-catenin is structurally related to vinculin. X represents uncharacterized attachment proteins involved in coupling cadherins to actin filaments.(Font: Alberts et al., 1993)
A més de les cadherines, molècules d’adhesió cel·lular de la superfamília de les inmunoglobulines participen en les unions d’ancoratge, però no formen complexes d’unió Figure 22-3. Three of the N-CAMs produced by alternative splicing of the primary transcript produced from the single N-CAM gene. N-CAM 180 (180,000 MW) and N-CAM 140 are anchored in the membrane by a single hydrophobic α helix and differ in the length of their cytoplasmic domains. N-CAM 120 is attached to the membrane by a glycosylphosphatidylinositol (GPI) anchor. Each of these three N-CAMs can also vary in the length of the poly α (2→8) sialic acid chain, whose attachment site is indicated. [After G. M. Edelman, 1988, Biochemistry27:3533; and T. M. Jessell, 1988, Neuron1:3.] (Font: Lodish et al., 2000)
Altres tipus de molècules d’adhesió cel·lular, com les selectines participen en la formació d’unions d’ancoratge de caràcter temporal Figure 10-42. The protein-carbohydrate interaction that initiates the transient adhesion of neutrophils to endothelial cells at sites of inflammation. (A) The lectin domain of P-selectin binds to the specific oligosaccharide shown in (B), which is present on both cell-surface glycoprotein and glycolipid molecules. The lectin domain of the selectins is homologous to lectin domains found on many other carbohydrate-binding proteins in animals; because the binding to their specific sugar ligand requires extracellular Ca2+, they are called C-type lectins. A three-dimensional structure of one of these lectin domains, determined by x-ray crystallography, is shown in (C); its bound sugar is colored blue. Gal = galactose; GlcNAc = N-acetylglucosamine; Fuc = fucose; NANA = sialic acid.(Font: Alberts et al., 1993)
Un exemple d’adhesió cel·lular temporal és la unió dels leucòcits a les cèl·lules endotelials en resposta a senyals d’infecció o inflamació. Estes unions es deuen a selectines i a un tipus especial d’integrines Figure 22-4. Interactions between cell-adhesion molecules during the initial binding and tight binding of T cells, a kind of leukocyte, to activation endothelial cells. Once a T cell has firmly adhered to the endothelium, it can move (extravasate) into the underlying tissue. Activation of the endothelium requires signals, such as platelet-activating factor (PAF), that are released in areas of infection or inflammation; thus extravasation occurs only in such areas. [Adapted from R. O. Hynes and A. Lander, 1992, Cell68:303.] (Font: Lodish et al., 2000)
Les unions d’oclusió formen una barrera de permeabilitat a través dels epitelis Figure 19-2. The role of tight junctions in transcellular transport. Transport proteins are confined to different regions of the plasma membrane in epithelial cells of the small intestine. This segregation permits a vectorial transfer of nutrients across the epithelial sheet from the gut lumen to the blood. In the example shown, glucose is actively transported into the cell by Na+-driven glucose symports at the apical surface, and it diffuses out of the cell by facilitated diffusion mediated by glucose carriers in the basolateral membrane. Tight junctions are thought to confine the transport proteins to their appropriate membrane domains by acting as diffusion barriers within the lipid bilayer of the plasma membrane; these junctions also block the backflow of glucose from the basal side of the epithelium into the gut lumen. (Font: Alberts et al., 1993)
Estructura de les unions d’oclusió (c) A model showing how a tight junction might be formed by linkage of rows of protein particles in adjacent cells. [Part (a) from P. A. Cross and K. L. Mercer, 1993, Cell and Tissue Ultrastructure, A Functional Perspective, W. H. Freeman and Company, p. 50; part (b) courtesy of L. A. Staehelin; part (c) adapted from L. A. Staehelin and B. E. Hull, 1978, Sci. Am. 238(5):140, and D. Goodenough, 1999, Proc. Natl. Acad. Sci. USA 96:319.] (Font: Lodish et al., 2000) Figure 15-28. Tight junctions. (a) Thin-section electron micrograph of the apical region of two liver epithelial cells, illustrating the tight junction just below the microvilli and the adherens junction. From the apical region of these liver cells, which faces the lumen of the bile duct, phospholipids and other components of bile are secreted into the duct. (b) Freeze-fracture electron micrograph of a tight junction between two intestinal epithelial cells. The fracture plane passes through the plasma membrane of one of the two adjacent cells. The honeycomblike network of ridges of particles below the microvilli forms the tight junction.
Les unions comunicants permeten el pas de xicotetes molècules d’una cèl·lula a l’altra Figure 21-35. An electric synapse. (a) The plasma membranes of the presynaptic and postsynaptic cells are linked by gap junctions. Flow of ions through these channels allows electric impulses to be transmitted directly from one cell to the other. (b) Negatively stained, electron microscopic image of the cytosolic face of a region of plasma membrane enriched in gap junctions; each “doughnut” forms a channel connecting two cells. [Part (b) courtesy of N. Gilula.] Figure 22-8. Structure of gap junctions. (a) In this model, a gap junction is a cluster of channels between two plasma membranes that are separated by a gap of about 2 – 3 nm. (b) Both membranes contain connexon hemichannels, cylinders of six dumbbell-shaped connexin subunits. (c) Each connexin subunit has four transmembrane α helices. Two connexons join in the gap between the cells to form a gap-junction channel, 1.5 – 2.0 nm in diameter, that connects the cytoplasm of the two cells. (Font: Lodish et al., 2000)
Resumen dels mecanismes d’adhesió cèl·lula-cèl·lula i cèl·lula-matriu extracel·lular Figure 19-28. A summary of the junctional and nonjunctional adhesive mechanisms used by animal cells in binding to one another and to the extracellular matrix. The junctional mechanisms are shown in epithelial cells, while the nonjunctional mechanisms are shown in nonepithelial cells. A junctional interaction is operationally defined as one that can be seen as a specialized region of contact by conventional and/or freeze-fracture electron microscopy. Note that the integrins and cadherins are involved in both nonjunctional and junctional cell-cell (cadherins) and cell-matrix (integrins) contacts. The cadherins generally mediate homophilic interactions, whereas the integrins mediate heterophilic interactions. Both the cadherins and integrins act as transmembrane linkers and depend on extracellular divalent cations to function; for this reason, most cell-cell and cell-matrix contacts are divalent-cation-dependent. The selectins and integrins can also act as heterophilic cell-cell adhesion molecules: the selectins bind to carbohydrate, while the cell-binding integrins bind to members of the immunoglobulin superfamily. (Font: Alberts et al., 1993)











