Conclusions
Manganese deficiency decreases pecan leaf photosynthesis by reducing the number of chloroplasts per unit leaf area. Fernando S. Henriques * and Renato R. P. Coelho **. * Faculdade de Ciências e Tecnologia - Universidade Nova de Lisboa * *I.C.A.M. - Universidade de Évora. The Problem


Conclusions
E N D
Presentation Transcript
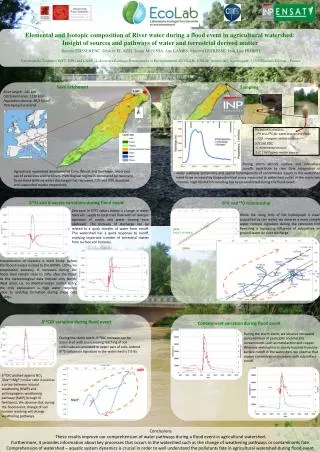
Manganese deficiency decreases pecan leaf photosynthesis by reducing the number of chloroplasts per unit leaf area Fernando S. Henriques* and Renato R. P. Coelho** *Faculdade de Ciências e Tecnologia - Universidade Nova deLisboa **I.C.A.M. - Universidade de Évora The Problem It has long been established that more than 80% of all the manganese contained in the chloroplasts is associated with PSII-enriched particles (Anderson et al., 1964), and various authors have shown that a tetranuclear Mn-center is required for the photoinduced water oxidation carried out at this photosystem (Yachandra et al., 1996). Such findings led to a generally accepted view that Mn deprivation should affect the photosynthetic process primarily by inhibiting electron donation at the PSII site and the subsequent electron transfer reactions. Reports by the overwhelming majority of authors supported this assumption. However, already in 1967, Peter Homann reported that leaves of Cassia obtusifolia responded to Mn deficiency in a manner different from all the other plant species he studied, displaying leaf photosynthetic rates on a chlorophyll basis higher than the controls, while their isolated chloroplasts exhibited Hill reaction rates comparable to or even higher than control chloroplasts. To explain these divergent results with Cassia, Homann proposed that “ higher plants could respond to Mn deficiency either by adjusting the number of chloroplasts per cell to the limited Mn supply or by forming disorganized chloroplasts with low chlorophyll content”. The first alternative is very interesting evolutively, but has received no further experimental support and has, at times, been looked at with some mistrust. This contradiction of Homann’s results with most published data led us to test the response to Mn deficiency of other plant species and we report here that pecan (Carya illinoinensis Koch, cv. Kiowa) leaves behave in a manner identical to that described for Cassia obtusifolia. Results and Discussion Data in Table 1 show that chl content decreases with increasing leaf Mn concentration, reaching only about 1/3 of the controls in the most severe case of Mn deficiency. Chl a:b and chl:car ratios remain essentially similar to controls, suggesting no major change in size or composition of PSII units. Table 1. Mn content in pecan leaves and its effects on leaf total pigment levels and pigment relative ratios.a Table 2. Mn effects on CO2 assimilation rate, stomatal conductance and intercellular CO2 concentration of pecan leavesa. Data in parentheses refer to photosynthetic rates per unit chl weight (mol mg -1chl hr-1) Variable Treatment Variable Treatment Control Mild def. Sev. def. Control Mild def. Sev. def. Mn content (mg g-1 dry wt) 795 112 72 CO2 assimilation rate (mmol m-2 s-1) 16.71.0a (14318) 9.30.9 (13423) 6.20.8 (14827) Chl content (mg cm-2 ) 423 254 152 Chl a:b ratio 4.00.1 3.70.1 3.50.1 Stomatal conductance (mol m-2s-1) 0.360.05 0.320.03 0.310.02 Carotenoid content (mg cm-2 ) 9.90.3 6.10.2 4.20.3 Intercellular CO2 concentration (mmol mol-1) 27315 30313 31510 Chl: car ratio 4.2 4.1 3.7 a Results are the means SD of 7 separate measurements a Data are the means SD of 7 separate experiments; measurements were carried out at an irradiance of 150080mol m-2s-1and at ambient CO2 concentrations. Table 3. The slight decrease ( 10%) of the Fv/Fm ratio with increasing Mn deficiency indicates that the intrinsic yield of PS II photochemistry was not reduced, i. e., that there is no decrease in the photochemical capacity of the PS II units remaining in Mn-deficient leaves. The time taken to rise chl fluorescence from its minimum to maximum level remains essentially unchanged among treatments, confirming data on pigment ratios that no significant changes have occurred in the relative sizes of the antenna : reaction center ratios. Finally, the measured decrease of PQ pool sizes with increasing Mn deficiency indicates a reduction in the number of PS II units per unit leaf area. The observation that the photosynthetic rates presented in Table 2 are linearly correlated with these PQ pool sizes further confirm that this pool of electron acceptors are fully involved in CO2 assimilation and not associated with inactive PS II units. Fig.1. Electron micrograph of chloroplasts from a Mn-deficient pecan leaf Table 3. In vivochlorophyll a fluorescence parameters in control and Mn- deficient pecan leavesa Variable Treatment Control Mild def. Sev. def. FV/FM 0.83 (0.01) 0.79 (0.01) 0.76 (0.01) TM 698 (24) 722 (27) 716 (29) PQ pool sizeb 841 (40) 526 (30) 318 (27) a Results are expressed in arbitrary units and refer to one specific measurement; in parentheses are the standard deviations of 10 separate experiments b PQ pool size was estimated from the area above the fluorescence induction curve Conclusions From the data gathered, it is concluded that Mn deficiency depresses the leaf photosynthetic capacity primarily by reducing the number of PS II units per unit leaf area and that those remaining preserve their full photochemical capacity. Also, given that no reduction in number or size of thylakoidal lamellae was observed in chloroplasts of Mn-deficient leaves, it is further concluded that the reduction of PS II units is accounted for by a reduction in the number of chloroplasts per unit leaf area, rather than by a decrease in the number of PSII units per individual chloroplast. References Anderson, J., Boardman, N. and D. David, 1964. Biochem. Biophys. Res. Comm. 17: 685. Govindjee, 1995. Aust. J. Plant Phys. 22: 131. P. Homann, 1967. Plant Physiol. 42: 997. Yachandra, V., Sauer, K.,and M. klein, 1996. Chem. Rev. 96: 2927. The approach to the problem The reduction in CO2 assimilation capacity observed in Mn-deficient leaves results primarily from a reduction in total PSII activity, although the exact mechanism has not been fully clarified and may eventually turn out to be variable with the plant species. To determine whether in pecan leaves this reduction in net photosynthesis per unit leaf area is due to an impairment of the PSII units present in the Mn-deficient leaves, to a reduction in the number of total photosynthetic units while the remaining ones preserve their functional capacity, or to both, we resorted mostly to the technique of chlorophyll a fluorescence induction (FI) in dark-adapted leaves. Chlorophyll fluorescence at room temperature emanates mainly from PSII and thus this technique provides direct information on this photosystem’s performance (Govindjee, 1995). For our present purposes, we need only to discuss here the ratio of variable to maximum (Fv/Fm) fluorescence which is indicative of the intrinsic yield of PSII photochemistry, the rise time to reach the maximum level of fluorescence (Tm), and the area over the fluorescence curve which is proportional to the PQ pool size associated with photosystem II. If the PSII units present in Mn-deficient leaves show reduced Fv/Fm ratios relative to controls, then the maximum functional capacity of such units is lowered and this must account, at least partly, by the decreased rates of net photosynthesis measured in these leaves. On the other hand, if the Fv/Fm ratio undergoes no significant reduction with Mn deficiency, then this is an indication that no damage has occurred to the reaction centers of PSII and the lowered net photosynthetic rates measured in Mn-deficient leaves must result from a reduction in the number of PSII units per unit leaf area. We also used electron transmission microscopy to search for ultrastructural changes in chloroplasts of Mn-deficient leaves. Table 2 shows that CO2 assimilation rates of Mn-deficient leaves are much lower than the controls on a leaf area basis, but are identical when expressed per unit chl. Decreased CO2 availability at the carboxylation sites can be ruled out as a possible cause for such lowering, as no reduction in stomatal conductance is observed in Mn-deficient leaves and intracellular CO2 concentrations remain high. Transmission electron microscopy showed that chloroplasts from Mn-deficient leaves are indistinguishable from controls, displaying well differentiated grana and stroma lamellae regions. No change in thylakoid’s organization or development was observed relative to controls.