Download

1 / 75
780 likes | 1.31k Views
Alternative Splicing Hedi Hegyi, PhD @ Institute of Enzymology, Budapest http://www.enzim.hu/~hegyi/ Szeged University, Biochemistry Course Oct 31, 2007. Scientific American 2005/04. Scientific American 2005/04.

E N D
Alternative SplicingHedi Hegyi, PhD@ Institute of Enzymology, Budapesthttp://www.enzim.hu/~hegyi/Szeged University, Biochemistry Course Oct 31, 2007
Scientific American 2005/04 • Spring of 2000. Molecular biologists placing dollar bets: how many genes in human genome? • 90,000? 153,000? C.elegans:19,500, Maize:40,000 • 35,000, 30,000, • a paltry 25,000!
C-value paradox: Complexity does not correlate with genome size. (C.A. Thomas, Jr, 1971) 6.7 x 1011 bp Amoeba dubia 3.4 x 109 bp Homo sapiens
N-value paradox: Complexity does not correlate with gene number. ~25,000 genes ~26,000 genes ~50,000 genes
Discovery of Alternative Splicing • First predicted by Walter Gilbert in 1978 • - First discovered for an Immunoglobulin heavy chain gene in 1980 (Edmund Choi, Michael Kuehl & Randolph Wall, Nature286, 776 - 779) • - Alternative splicing gives two forms of the protein with different C-termini: • - 1 form is shorter and secreted • - Other stays anchored in the plasma membrane via C-terminus
Alternative splicing of the mouse immunoglobulin μ heavy chain gene S - signal peptide Red – untranslated region V - variable region Green – membrane anchor C - constant region Yellow – end of coding reg. for secreted form
Splicing & the spliceosome Structure • 60S dynamic structure – a large complex consisting of ~ 150 proteins • Five small nuclear RNAs (U1, U2, U4, U5 & U6) • RNAs assemble with proteins to form snRNPs (“snurps”) • Protein splicing factors Assembly of spliceosome requires ATP Splicing defects • Estimation: 15% of all genetic diseases associated with mutated splice sites Green globule: RNA pol Yellow globule: spliceosome
Secondary structure of snRNAs U5 U1 U2 Orange - interaction with 5’ splice site Green – Interaction with branch site Blue - interaction between U2 and U6 Tan - Sm-binding site (PuAU4-6GPu) flanked by two stem-loop structures U6 U4
U1 snRNA • Contains conserved sequence complementary to 5’ splice site of nuclear mRNA introns • Contains pseudouridine(y) 5’ splice site Upstream exon GUAAGU-------3’ :::::: 3’---CAUUCA---cap-5’ U1 snRNA
Splice-site recognition 5’ splice site 3’ splice site upstream exon downstream exon branch site ---AGGUAAGU-----------A--------(Py)nNCAGG ~ 20 – 50 nts Intron Branch site in yeast: often 5’- UACUAAC-3’
Splice Site Conservation Splice Junction 5’ XX YY 3’ E I E I E Donor (5’) SS Acceptor (3’) SS
Splice Site Conservation Splice Junction 5’ XX YY 3’ E I E I E Donor (5’) SS Acceptor (3’) SS
U5 U4 U6 ATP AG U1 U5 U2 U4 U6 Splicing mechanism 5’ splice site branch site 3’ splice site Exon 1 Intron Exon 2 U1 U2 GU A AG
Factors Playing a Role in Exon Recognition 1. Evolution appears to have weakened splice sites Derived from 253, only 3% of the S. cerevisiae genes contain introns No Alternative Splicing Derived from 4,697 S. pombe genes; approximately 43% of all genes contain introns Intron Retention Derived from 49,778, nearly 100% contain introns 75% Alternative Splicing - GTCCATTCA - 5' U1
Factors Playing a Role in Exon Recognition Exon Recognition is complex Exon Definition Intron Definition Complexity means multiple points of possible regulation and that exons could be skipped by failing to get all the pieces in place The ability to form or disrupt these interactions is thought to play a key role in alternative splicing!!! Nature, Vol. 418, p. 236, 2002
Intron statistics SpeciesAverage Average Average Average % exon exon No. intron No. length(kb) kb mRNA per gene Yeast 1 0 1.6 1.6 100 Nematode 4 3 4.0 3.0 75 Fruit fly 4 3 11.3 2.7 24 Chicken 9 8 13.9 2.4 17 Mammals 7 6 16.6 2.2 13 Human genes MedianMean Size of internal exons 122 bp 145 bp Number of exons 7 8.8 Size of introns 1023 bp 3356 bp
Recognizing Exons - Are Splice Sites Enough? Most mammalian genes contain more than one intron Extreme Examples: Collagen Gene - 50 exons and a 40 kb precursor RNA DMD Gene - 79 exons, 2.3 mb precursor, intron 20 is 180 kb Neurexin - has a 0.5 mb intron! Most genes are uninterrupted in yeast, but most genes are interrupted in flies and mammals
Support for the Exon Definition Model Exon size is conserved in vertebrates Intron size is not Recognition requires a consistent target size
Additional Cis Elements and Trans-Acting Factors - Splicing has Enhancers and Silencers - They function to modulate spliceosome formation Recruiting SR proteins can stabilize E complex formation Blocking snRNP or protein interactions can prevent E complex formation ESE - Exonic Splicing Enhancer ESS - Exonic Splicing Silencer ISE - Intronic Splicing Enhancer ISS - Intronic Splicing Silencer
Models of Exon Recognition - Final Points Trans-Factor Interaction with Exon differs from Interaction with Introns
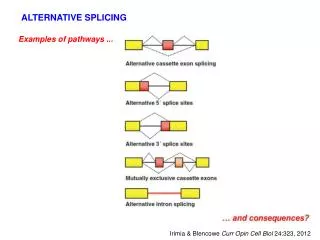
Types of Alternative Splicing 38% 18% 8% 3% Remaining 33% Cell 126:37 (2006) Nature Reviews: Genetics 5:773 (2004)
How Prevalent is Alternative Splicing? No one really knows for sure. EST Database estimates between 35 - 60% of protein coding gene have alternative mRNAs Caveat - These databases contain sequences derived from aberrant, as well as, alternative splicing, they are typically 3' and 5' end biased, and have insufficient number to infer frequency Therefore, database mining may overestimate the rate of alternative splicing
74% Array-Based Numbers Science 302, 2141-44 (2003)
Genome-Wide Survey of Human Alternative Pre-mRNA Splicing with Exon Junction Microarrays (Science, 2003) 10,000 multi-exon human genes in 52 tissues Conclusion: 74% of multi-exon human genes are alternatively spliced
Number of Splicing Isoforms per Gene by EST Comparison 3.8 Harrington et al. Nature Genetics 36:916 (2004)
Regulation of/by Alternative Splicing • Sex determination in Drosophila involves 3 regulatory genes that are differentially spliced in females versus males; 2 of them affect alternative splicing 1. Sxl (sex-lethal) - promotes alternative splicing of tra (exon 2 is skipped) and of its own (exon 3 is skipped) pre-mRNA 2. Tra – promotes alternative splicing of dsx (last 2 exons are excluded) 3. Dsx (double-sex) - Alternatively spliced form of dsx needed to maintain female state Fig. 14.38
Alternative splicing in Drosophila maintains the female state Alternative splicing Sxl and Tra are SR proteins Tra binds exon 4 in dsx mRNA causing it to be retained in mature mRNA.
Known Roles of Alternative SplicingStamm et al. Gene 344:1-20 (2005) • Introduction of stop codons - • 25-35% of alternative splicing events introduce stop codons that either function to produce truncated proteins or regulate mRNA stability through the nonsense mediated decay (NMD) pathway
Nonsense Mediated Decay • A surveillance mechanism that selectively degrades nonsense mRNAs • Regulates gene expression by alternative splicing • - Transcripts containing a PTC (premature termination codon) are degraded rapidly 1/3rd of alternative transcripts contain premature termination codons Brenner, SE et al, PNAS January 7, 2003 vol. 100 no. 1 189–192
Known Roles of Alternative SplicingStamm et al. Gene 344:1-20 (2005) • Add new protein parts - • 75% of alternative splicing involves the protein coding region, in addition to truncations you can change the overall protein sequence
Known Roles of Alternative SplicingStamm et al. Gene 344:1-20 (2005) • Consequences of new protein parts - • Alter protein binding properties, eg. receptor/ligand • Alter intracellular localization, eg. membrane insertion • Alter extracellular localization, eg. secretion • Alter enzymatic or signaling activities, eg. TK truncations • Alter protein stability, eg. inclusion of cleavage sites • Insertion of post-translation modification domains • Change ion channel properties eg. slo
Known Roles of Alternative SplicingStamm et al. Gene 344:1-20 (2005) • Coordinated Regulation of Biological Events • Potassium channel activity associated with hearing (slo) • Muscle contraction • Neurite (axon or dendrite) growth • Cell differentiation • Apoptosis • Neuron development (Dscam) (TIBS 31:581-588, 2006)
The Power of Alternative RNA Splicing Drosophila DSCAM gene codes for an axon guidance receptor The final mRNA chooses 24 exons from 115 possibilities (20 constitutive exons and 4 alternatively spliced ones) Ig Loop 7 Trans-membrane Ig Loop 3 Ig Loop 4 Exon 4 12 Alternatives Exon 6 48 Alternatives Exon 9 33 Alternatives Exon 17 2 Alternatives 12 X 48 X 33 X 2 Equals 38,016 Possible mRNAs Genome has only 14,800 genes!
Evolutionary Overview of Alternative Splicing • Introns unlikely to have been derived from ancient genes • Multi-intron genes probably predated alternative splicing • Most eukaryotes have introns but alternative splicing prevalent only in multicellular organism • S.cerevisiae has only 253 introns (3% of its genes) and only 6 genes have 2introns • S. pombe: 43% of its genes have introns (usually 40-75 nt) • S.cerevisiae and S. pombe have NO alternative splicing
Finding Alternatively Spliced Exons • Compare cDNA & genomic DNA sequences • Compare ESTs & genomic DNA sequences • Compare protein & genomic sequences
Large-scale multiple alignment of expressed sequences • Databases: • tens of thousands of mRNAs • millions of ESTs • From large-scale alignments: 60-80% of all human genes undergo alternative splicing.
Alu elements • Length = ~300 bp • Repetitive: > 1,400,000 times in the human genome • Constitute >10% of the human genome • Found mostly in intergenic regions and introns • Propagate in the genome through retroposition (RNA intermediates).
Alu elements can be divided into subfamilies The subfamilies are distinguished by ~16 diagnostic positions.
Alu-containing exons • Out of 1,182 alternatively spliced cassette exons, 62 have a significant hit to an Alu sequence. • Out of 4,151 constitutively spliced exons, none has a significant hit to an Alu sequence. all Alu-containing exons are alternatively spliced. Graur et al., Genome Res. (2002)
The minus strand of Alu elements contains “near” splice sites • The minus strand of Alu contains ~3 sites that resemble the acceptor recognition site: Consensus acceptor site:YYYYYYNCAG/R Alu-J: (127-114) :TTTTTTGtAG/A • The minus strand of Alu contains ~9 sites thatresemble the consensus donor site: Consensus donor site: CAG/GTRAGT Alu-J: (25-17) : CAG/GTGtGA
Alu exonization The selection of AGs in the 3SSs of Alu-derived exons are underlined 3 genetic diseases: 1. COL4A3- Alport syndrome 2. GUSB- Sly syndrome 3. OAT- OAT deficiency Lev-Maor G, et al, Science, 2003.
Factors Playing a Role in Exon Recognition - Sequence features of alternatively regulated exons are different from constitutive exons. - These features are conserved between species. Sorek, Genome Res 14:1617 (2004)
Large-scale identification of alternative splice variants of human gene transcripts using 56,419 cDNAs Distribution of the length difference between the alternative splicing variants Takeda, J.-i. et al. Nucl. Acids Res. 2006 34:3917-3928; doi:10.1093/nar/gkl507
Large-scale identification of human alternative splice variants (A)‘motif-changed’ (B)‘subcellular localization-changed’ (C)‘transmembrane domain-changed’
Alternative splicing databases (1,560,000 hits in google) • - Alternative Splicing & Transcript Diversity DbASTD • http://www.ebi.ac.uk/astd/ • - SpliceMiner (querying EVDB - Evidence Viewer Database) • http://discover.nci.nih.gov/spliceminer/ • Hollywood • http://hollywood.mit.edu • -Human Alternative Splicing Db (HASDB), • http://www.bioinformatics.ucla.edu/~splice/HASDB/ • Putative Alternative Splicing Database, • PALS db, http://palsdb.ym.edu.tw/
Structure of ASTD databases are integrated, cross-linked and are available through a variety of interface tools ASTD data are integrated with Ensembl genome annotation Stamm, S. et al. Nucl. Acids Res. 2006 34:D46-D55
Spliceminer (NCBI) Querying EVDB (Evidence Viewer DB). Composite of five separate interactive queries. Each query corresponds to a different Affymetrix HG-U133A Probe. The composite permitsfacile comparison of the exons that are targeted by each of the probes. For example, the probes for exons 16 and 18 uniquely identify the splice variants NM_006487 and NM_006485, respectively.
Evolutionarily conserved and diverged alternative splicing events show different expression and functional profiles (Kan, NAR, 2005) Kan, Z. et al. Nucl. Acids Res. 2005 33:5659-5666; doi:10.1093/nar/gki834