Download

1 / 24
250 likes | 468 Views
Chromozómy a chromatin. strukturní uspořádání genetické informace. Chromozómy. NOR: 18S- 5,8S- 26S rDNA. Karyotyp – počet, typy a velikosti chromozómů. Klasický karyotyp (metafáze). Flow karyotyp (značení FISH). Doležel et al. 1999. Telomery.

E N D
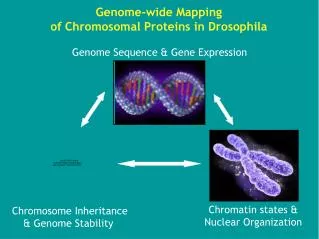
Chromozómy a chromatin strukturní uspořádání genetické informace
Chromozómy NOR: 18S- 5,8S- 26S rDNA
Karyotyp – počet, typy a velikosti chromozómů Klasický karyotyp (metafáze) Flow karyotyp (značení FISH) Doležel et al. 1999
Telomery DNA/proteinové struktury zajišťující stabilitu chromozómových konců Repetitivní sekvence přidávaná telomerázou po replikaci (TTTAGGG)n u Arabidopsis U některých rostlin i typická savčí sekvence: (TTAGGG)n
Udržování délky telomer telomeráza: prodlužuje konce chromozómů dle RNA templátu - počet repetic se liší u druhů, vývojových stádií, typu buněk, ale i jednotlivých chromozómů
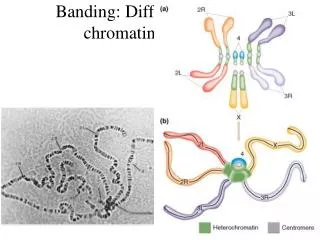
Centromery - definovány přítomností CenH3 Typy: - bodové (125 bp, kvasinka Saccharomyces) - holocentrické chromozómy (Cen H3 všude, př. Luzula – umožňuje snadnou fragmentaci) - klasické – různě rozsáhlá oblast heterochromatinu (repetice, TE) - dány epigeneticky (neocentromery) - různá síla u hybridů ((telocentrické, akrocentrické, metacentrické, submetacentrické))
Modifikací cetromerového histonu CenH3 u Arabidopsis lze získat haploidní potomstvo Ravi and Chan 2010
Chromozomální teritoria oblast jádra, kde se nalézá určitý chromozóm (po mitóze dekondenzace cca 2 hod, 2,5 x zvětšení) Rablovo uspořádání chromozómů centromery a telomery na opačných koncích (u velikosti nad cca 500 Mb)
Experimentální doložení existence chromozomálních teritórií - značení chromozómů či jejich částí pomocí FISH poškození laserem a detekce reparace Cremer and Cremer 2010
Pohlavní chromozómy u rostlin - chromozómy zodpovědné za determinaci pohlaví - zodpovědný může být jeden či větší množství lokusů • U rostlin se pohlavnost vyvinula nezávisle a opakovaně u různých taxonů (celkově 5% druhů, u 75% čeledí) • - játrovky, jinan, silenka, šťovík, chmel, topol, … • Morfologické členění pohlavních chromosómů: • homomorfní • heteromorfní • - polymorfní - více než dva typy • Rumex acetosa: male XY1Y2, female XX • Humulus lupulus var. cordiflorus: male X1X2Y1Y2, female X1X1X2X2
Evoluce pohlavních chromozómů Podmínka vzniku – vznik mutací vedoucích k samčí a samičí sterilitě u dvou lokusů v poměrně silné vazbě (mezistupeň gynodioecie) (model samice XX – X nese nefunkční alelu genu nutnou pro vývoj samčích pohlavních orgánů samec XY – Y nese gen potlačující tvorbu samičích orgánů ve vazbě s funkčí alelou pro tvorbu samčích) Evolučně mladé – homomorfní (rekombinace omezena jen na části) Degenerací (inverze, kumulace repetitivních sekvencí a TE, delece) vznikají heteromorfní – zprvu nárůst, pak redukce Rozpadem (translokacemi) mohou vznikat polymorfní Polyploidie znesnadňuje vytvoření dvoudomosti! U pohlavních chromozómů chybí některé TE!
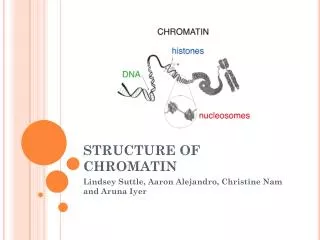
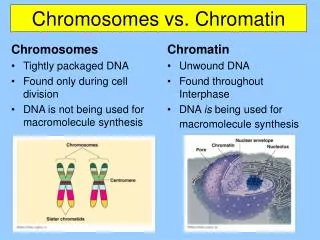
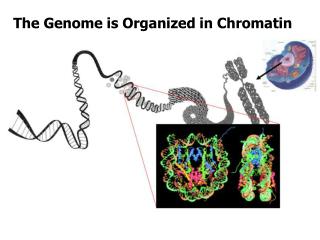
Chromatin (Walther Flemming 1882) = DNA + asociované proteiny euchromatin x heterochromatin heterochromatin fakultativní x konstitutivní
Chromatin (DNA + asociované proteiny) Genetická informace = sekvence DNA - regulační, protein-kódující, RNA-kódující Epigenetická informace - přístupnost interagujících proteinů, transkripční aktivita transkripčně aktivní transkripčně neaktivní rozvolněný, přístupný kompaktní, nepřístupný
Epigenetické modifikace chromatinu - mitoticky a/nebo meioticky dědičné změny v aktivitě genů - nemohou být vysvětleny změnami v primární sekvenci DNA - kovalentní úpravy chromatinu neměnící sekvenci DNA - metylace DNA - posttranslační modifikace histonů VZÁJEMNÁ PROVÁZANOST! - typy histonů euchromatin heterochromatin
Nukleozóm oktamer histonů (malé bazické proteiny): 2 x H2A, 2 x H2B, (2 x H3, 2 x H4) + 147 bp DNA VARIABILNÍ SLOŽENÍ: - varianty jednotlivých histonů (CenH3, H3.3, H2A.Z, …) - posttranslační modifikace histonů
Struktura 30nm vlákna Solenoid nebo ZigZag? - stále nejasné solenoid Li and Reinberg 2011
Vyšší strukturní úrovně chromatinu • smyčky s aktivně transkribovanými geny • nedostatečně prozkoumané • MARs (matrix attachment regions) • také SARs (scaffold attachment reg.) • ukotvují oblasti kolem kódujících oblastí (funkčních genů) k bílkovinné matrix chromozómu nebo jaderné matrix interfázního jádra • AT bohaté, kolokalizují s „izolátory“ • (sekvence bránící šíření heterochromatinu) • vzdáleny vzájemně 3kb až 100kb • LBARs • (loop basement attachment regions) • - organizují genom do velkých smyček (vzdálenost 20kb až 100kb)
Propojení nukleozómů - délka spojovací sekvence mezi nukleozómy (násobky 10ti bp – 20 až 90 bp) - průměrné délky druhově, ale i pletivově specifické (20 bp kvasinka, 30 Arabidopsis, 40 savci) - celkem úseky 167 – 237 bp dlouhé ( časté délky repetic)
Propojení nukleozómů Interakce mezi nukleozómy - N-konce H4 interagující s H2A.H2B těly v nukleozómech paralelního vlákna - přítomnost H2A.Z zřejmě brání paralelní interakci vláken Linker histon H1 - má zásadité amino i karboxy-konce a interaguje s histony i s DNA - stabilizace vyšších struktur (30nm), fosforylace během buněčného cyklu) - vliv délky spojovací sekvence: delší - potřebují H1 a bývají kompaktnější – heterochromatin kratší – menší vliv H1, volnější – aktivnější chromatin
Poloha nukleozómů • umístění nukleozómů na DNA není nahodilé (ale je měnitelné) • - sekvence DNA • - metylace DNA • - modifikace/typy histonů • - transkripce DNA • regulace/modulace transkripce • - „unstable nucleosom region“ (dříve „nucleosom-free region“) před počátkem transkripce (především konstitutivně exprimované geny) - nestabilní nukleozómy s H3.3 a H2A.Z histony • - obklopené stabilně umístěnými nukleozómy s histonem H2A.Z • - nukleozómy rovněž pomáhají definovat exony (centrální umístění, i bez probíhající transkripce!)
Histonový kód • - kovalentní posttranslační modifikace (PTM) • - modifikace především N-konců (do prostoru) • vysoká komplexita • „epigenetické instrukce“ k nakládání s DNA
Některé PTM histonů se vzájemně ovlivňují a mají vícečetné role např. fosforylace H2A – značení poškozené DNA, ale i role při regulaci transkripce a spiralizaci, PCD Rossetto et al. 2012
Histonový kód Fosforylace – zřejmě především transientní značka, různé role Acetylace – především „výkonná modifikace“ oslabení vazby s DNA - K-Ac - vazba proteinů s bromodoménami Metylace – signální role ( stabilní), represní i aktivační značka (~ pozici) - K-Me – vazba chromodomén, TUDOR-like domén - klíčová role v regulaci metylace DNA a aktivity chromatinu - H3K9me2 (dimetylace lyzinu na pozici 9 v histonu H3) - klíčová represní značka heterochromatinu - H3K4me3 – aktivační značka chromatinu - H3K27me3 – represní značka u vývojově regulovaných genů
Reprodukce nukleozómů po replikaci- histonové tetramery (H3/H4) a dimery (H2A.H2B) se nedělí mezi dceřinná vlákna (epigenetická informace přenášena jinak!)- přenos parentálních (Asf1) de novo depozice (CAF1, Asf1) - H2A.H2B doplňovány i s prodlevou (mimo replikaci)