Download

1 / 42
420 likes | 584 Views
From the Proliferating Microsphere to the Chemoton. Chrisantha Fernando School of Computer Science University of Birmingham, UK San Sebastian, September 2006. The Chemoton. Tibor Ganti’s hypothetical pre-enzymatic minimal unit of life (1971, 1973, etc..)

E N D
From the Proliferating Microsphere to the Chemoton. Chrisantha Fernando School of Computer Science University of Birmingham, UK San Sebastian, September 2006.
The Chemoton • Tibor Ganti’s hypothetical pre-enzymatic minimal unit of life (1971, 1973, etc..) • Stoichometrically coupled autocatalytic metabolism, template and membrane systems.
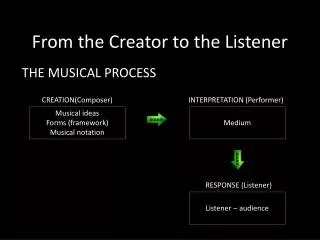
Templates control but don’t encode
The Origin of the Chemoton • Q1. How could the ‘notorious’ (L.Orgel) formose cycle metabolism be so nicely channeled? • Ganti proposed the proliferating microsphere as a chemoton ancestor. • A chemoton without templates, just metabolism and boundary.
Q2. Why would a proliferating microsphere become a chemoton? • Why would templates have interposed themselves to become rate-limiting in a previously well functioning microsphere? • What immediate selective advantage would the Ganti type (i.e. non-encoding) template replication system confer to a proliferating microsphere? ?
This is disturbing for the non-enzymatic chemoton idea. • Early template replication is likely to have been slow (low rates of p-bond formation) so the chemoton would be less fit than a proliferating microsphere! • If Ganti’s claim of a non-enzymatic chemoton is true, he cannot call upon the ribozyme functionality of templates. • The cause of evolution from PM to C is not explained by chemoton theory.
Possible Explanation 1 • If PM metabolism is initially inefficient at producing membrane elements, R. • If template polycondensation reactions increase the rate of R production. • Then there is selection for increased initiation and propagation rates of polycondensation. • This is best achieved by clean replication of templates with Tm = chemoton operating temperature, rather than messy, branched polymer formation, i.e. chemoton selection is against all kinds of product inhibition.
Possible Explanation 2 • Template Length can confer a weak Lamarkian kind of heredity. • Long templates favour slow replication with high resting metabolite concentrations. • Short templates favour faster replication. • In harsh conditions, templates elongate, and this makes chemoton offspring have higher metabolite concentrations.
Possible Explanation 3 • Short early templates conferred an advantage to the chemoton by having ribozyme effects. • Ganti was wrong about the possibility of a chemoton without encoded catalysis.
Q3.Do Long (non-enzymatic) templates make faster Chemotons? • Once a Chemoton had formed, could between-chemoton selection be a driving force for long template replication?
Not According to Ganti’s Model of the Chemoton. • According to Ganti’s model, long template replication ALWAYS results in slower chemotons, if template replication is rate limiting. • This effect can be counteracted but not reversed by increasing initiation and propagation rates. • But then templates loose their ‘control’ function.
The Unrealistic Model • Assumes that above a [monomer] threshold, strands denature, initiate, and propagate. • Actually, one sees dimer, trimer formation predominating at high monomer concentration, and elongation predominating at low monomer concentration. • Also, there is no threshold! • Also, sequence dependent stacking effects influence the capacity for replication, even in the absence of ribozyme effects.
Chaos, due to leftover monomers after division, if [V]* is high. So What? [V]* = 35.0
No Chaos Left : [V]* = 5.0 Right : Tsize reduced by 10x
High Propagation Rate Low Propagation Rate Mean [V’] [Template Concentration] Mean Period Length Length Initialize chemotons with the same initial MASS, i.e. for longer templates one must start with a lower template concentration!
A more realistic model suggests selection may exist for short template replication • A stochastic model of template-replication shows that elongation would have been a problem for template replication. • Using what was learnt from this model, when elongation is incorporated into a chemoton model, there is indeed selective pressure to convert elongating templates to replicating templates with Tm near the operating temperature of the chemoton.
A stochastic model helps us understand template dynamics. • I’m not going to go into the nitty gritty of this stochastic model, but will present the main findings. • Template elongation successfully scuppers template replication at low temperature. • At high temperature, oligomer replication results in a skew towards very short strands.
Eors’ ODE Model of Template Replication • Considers 3 strands each double the length of the previous strand, in a coupled replication chain.
[conc] Low Temp High Temp time
Low Temp, High [Monomer] Elongation dominates Low Temp, Low [Monomer] Elongation dominates Low Temp, Low [Monomer] E and G decay at equal rates. Elongation dominates
Adding this new template model to the chemoton model Short Strands Predominate
[a] [b] [c] [g] [e] Note elongating template concentration decreases because it cannot double per chemoton replication.
What happens to chemoton replication rate when we increase monomer incorporation rate? Log monomer incorporation rate No further influence since templates are no longer rate-limiting.
Conclusion • If R cannot be produced by other means, then more efficient template polycondensation would be selected for. • However, if R is produced by other means as in the PM, then template polycondensation rate only has a slight influence on chemoton replication rate, by reducing back-flow into metabolism of V’
Log monomer incorporation rate
Furthermore • All templates can be lost from the chemoton if they do not double within the time of the chemoton. • If stoichiometric coupling assumption is relaxed, then only the rapidly replicating templates can survive. • Therefore in transition from PM to C, only templates capable of rapid replication would have been selected for.
If templates do survive. • Then the extent of the benefit to the chemoton depends on the tendency for metabolism to run in reverse without templates absorbing V’. Reverse rate = 0.1 Reverse rate = 0.00001 vs.
Short template replication could have arisen in the chemoton (from elongation), if template polycondensation could reduce back reactions of metabolism.
Conclusion • In trying to understand why a PM would evolve into a C, one possibility is that messy polymerization reactions would have been present in the PM. • PMs with greater template replication capacity would have been more efficient because in such PMs either metabolism would have been more irreversible, or R production would have been increased.
Thanks to • Eors Szathmary • Guenter Von Kiedrowski • Johan Elf and Mons Ehrenburg