Download
1 / 15
150 likes | 325 Views
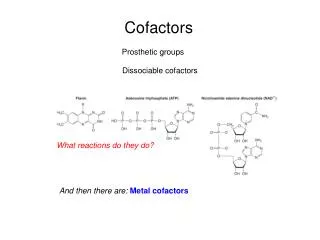
Cofactors. Prosthetic groups. Dissociable cofactors. What reactions do they do?. And then there are: Metal cofactors. Many Fe Cofactors. How much Iron in a Human? Typically in an adult human there is 4-5 grams of Fe comprising: hemoglobin - 2.6 g (65%) myglobin - 0.13 g (6%)

E N D
Cofactors Prosthetic groups Dissociable cofactors What reactions do they do? And then there are: Metal cofactors
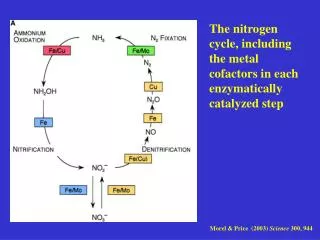
Many Fe Cofactors How much Iron in a Human? Typically in an adult human there is 4-5 grams of Fe comprising: hemoglobin - 2.6 g (65%) myglobin - 0.13 g (6%) transferrin - 0.007 g (0.2%) ferritin - 0.52 g (13%) hemosiderin - 0.50 g (12%) catalase - 0.004 g (0.1%) cytochrome c - 0.004 g (0.1%) other enzymes - 0.14 g (3.6%)
Ancient Fe Cofactors Rieske Fe-S cluster Plant Ferredoxin “3 Iron” clusters Ferredoxin (cubanes) Rubredoxin
Rubredoxin: one redox Plant Ferredoxin : two redox 3-Fe : three redox cubane : four redox Fe(3+)Fe(3+) Fe(2+)Fe(3+) Fe(2+)Fe(2+) 1Fe(2+) 3Fe(3+) 2Fe(2+) 2Fe(3+) 3Fe(2+)Fe(3+) All Fe(2+)
Tetrapyrrole Biosynthetic Pathways Form’n rate limiting dimerizes x 4 via linear tetrapyrrole corrin + Fe RED w/o Fe P = propionate; A = acetate
Errors cause Porphryias: Either excess ALA Or Slow Uro’gen III - 6 - -6 H+
The porphyrias: clinical presentation, diagnosis and treatment European Journal of Dermatology. Volume 16, Number 3, 230-40, May-June 2006, Author(s) : P Poblete-Gutiérrez, T Wiederholt, HF Merk, J Frank , Department of Dermatology, University Hospital Maastricht, The Netherlands, Department of Dermatology, University Hospital of the RWTH Aachen, Germany, Porphyria Center, University Hospital of the RWTH Aachen, Germany. Summary: The porphyrias comprise a clinically and genetically heterogeneous group of diseases mostly arising from a genetically determined dysfunction of specific enzymes along the pathway of heme biosynthesis. Based on the occurrence or absence of cutaneous symptoms and life-threatening acute neurological attacks, the different types of porphyrias can either be classified into cutaneous and non-cutaneous forms or acute and non-acute forms.
Heme Metabolism (because curious minds want to know) The largest repository of heme in the human body is in red blood cells, which have a life span of about 120 days. There is thus a turnover of about 6 g/day of hemoglobin, which presents 2 problems. First, the porphyrin ring is hydrophobic and must be solubilized to be excreted. Second, iron must be conserved for new heme synthesis. Heme is oxidized, with the heme ring being opened by the endoplasmic reticulum enzyme, heme oxygenase. The oxidation step requires heme as a substrate, and any hemin (Fe3+) is reduced to heme (Fe2+) prior to oxidation by heme oxygenase. The oxidation occurs on a specific carbon producing the linear tetrapyrrole biliverdin, ferric iron (Fe3+), and carbon monoxide (CO). This is the only reaction in the body that is known to produce CO. Most of the CO is excreted through the lungs, with the result that the CO content of expired air is a direct measure of the activity of heme oxygenase in an individual. In the next reaction a second bridging methylene (between rings III and IV) is reduced by biliverdin reductase, producing bilirubin. Bilirubin is significantly less extensively conjugated than biliverdin causing a change in the color of the molecule from blue-green (biliverdin) to yellow-red (bilirubin). The latter catabolic changes in the structure of tetrapyrroles are responsible for the progressive changes in color of a hematoma, or bruise, in which the damaged tissue changes its color from an initial dark blue to a red-yellow and finally to a yellow color before all the pigment is transported out of the affected tissue.
Extended Conjugation of Heme causes intense Soret band in visible.
Synthesis of Porphobilinogen and Heme The first reaction in heme biosynthesis takes place in the mitochondrion and involves the condensation of 1 glycine and 1 succinylCoA by the pyridoxal phosphate-containing enzyme, d-aminolevulinic acid synthase (ALA synthase). This reaction is both the rate-limiting reaction of heme biosynthesis, and the most highly regulated reaction. Mitochondrial d-aminolevulinic acid (ALA) is transported to the cytosol, where ALA dehydratase (also called porphobilinogen synthase or hydroxymethylbilane synthase) dimerizes 2 molecules of ALA to produce the pyrrole ring compound porphobilinogen. The next step in the pathway involves the head-to-tail condensation of 4 molecules of porphobilinogen to produce the linear tetrapyrrole intermediate, hydroxymethylbilane. This enzyme is called uroporphyrinogen I synthase. Hydroxymethylbilane has two main fates. The most important is regulated, enzymatic conversion to uroporphyrinogen III, the next intermediate on the path to heme. Genetic defects that cause increased ALA synthase activity or decreased uroporphyrinogen I synthase activity lead to the disease known as acute intermittent porphyria, which is diagnosed by the excretion of excess porphobilinogen (a condition that is not obvious from the color of the urine). In the cytosol, the acetate substituents of uroporphyrinogen (normal uroporphyrinogen III or abnormal uroporphyrinogen I) are all decarboxylated by the enzyme uroporphyrinogen decarboxylase. The resultant products have methyl groups in place of acetate and are known as coproporphyrinogens, with coproporphyrinogen III being the , important, normal intermediate in heme synthesis. Coproporphyrinogen III is transported to the interior of the mitochondrion, where 2 propionate residues are decarboxylated, yielding vinyl substituents on the 2 pyrrole rings. The colorless product is protoporphyrinogen IX. In the mitochondrion, protoporphyrinogen IX is converted to protoporphyrin IX (structure shown below) by protoporphyrinogen IX oxidase. The oxidase reaction requires molecular oxygen and results in the loss of 6 protons and 6 electrons, yielding a completely conjugated ring system, which is responsible for the characteristic red color to hemes. The final reaction in heme synthesis also takes place in the mitochondrion and involves the insertion of the iron atom into the ring system generating heme b. The enzyme catalyzing this reaction is known as ferrochelatase.
The Pyranopterin Cofactor: oldest organometallic cofactor? in molybdenum enzymes in tungsten enzymes
Moco Biosynthesis We know a lot about Moco biosynthesis Mendel 2005
Another sulfido iron molybdenum cofactor: FeMoco in N2ase We don’t know what ‘X’ is yet… X Another di-metallic cofactor: NiFe cluster in H2ase
FeFe cluster in H2ase Note Fe4S4 cluster NiFe cluster in H2ase