Download

1 / 12

E N D
Chap. 6. Problem 1. After harvesting, the simple sugars present in the kernels of corn continue to be enzymatically converted into starch. This sugar has a very sweet taste, whereas starch does not. Thus, within a day or so of harvest, untreated corn no longer tastes as sweet. To preserve sweetness, freshly picked corn can be heated (blanched) for a few minutes to denature the enzymes that synthesize starch. This preserves the sweet flavor of fresh corn. During freezing, any remaining starch synthesis enzyme activity is minimal due to the low temperature of storage.
Chap. 6. Problem 4. The data are consistent with the possibility that the hexokinase-substrate complex is actually thermodynamically more stable than free hexokinase. The additional interactions formed between the enzyme and its substrate must contribute to the stabilization of hexokinase. Because the energy of the hexokinase-substrate complex is lower than free hexokinase, the activation energy barrier separating the folded and unfolded states of the enzyme is increased in the presence of substrate. This reduces the extent of denaturation that occurs when the substrate is present in solution with the enzyme.
Chap. 6. Problem 7. Enzymes are catalysts that change the rates of reactions without affecting reaction equilibria. Thus, (b), (e), and (g) are correct as all of these parameters are connected to the reaction rate. (d) is not correct as the energy of the transition state actually is decreased, not increased, by the enzyme due to the complementarity of its structure to the transition state. Because reaction equilibria, and the related term ∆G’0 for a reaction, are not affected by an enzyme, (a), (c), and (f) are not observed.
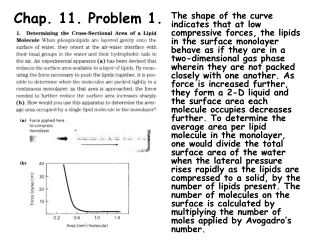
Chap. 6. Problem 8a. Part (a). In this problem, we are given the values of V0 (0.25 Vmax) and Km. On substitution of these parameters into the MM equation, we can solve for [S]. Namely, V0 = Vmax[S]/(Km +[S]) or 0.25Vmax = Vmax[S]/(Km + [S]) After canceling Vmax terms and rearranging the equation it becomes 0.25Km + 0.25[S] = [S] or 0.25Km = 0.75[S] On solving for [S] we get [S] = 0.33Km = (0.33)(0.005M) = 1.7 x 10-3 M. Part (b). See next slide.
Chap. 6. Problem 8b. Part (b). To determine the fraction of Vmax (V0/Vmax) that is obtained at any [S] relative to the Km, first rearrange the MM equation to the form V0/Vmax = [S]/(Km + [S]) And then substitute in [S] values expressed as a function of Km. 1) For [S] = 0.5Km V0/Vmax = 0.5Km/(Km + 0.5Km) = 0.5/1.5 = 0.33 2) For [S] = 2Km V0/Vmax = 2Km/(Km + 2Km) = 2/3 = 0.67 3) For [S] = 10Km V0/Vmax = 10Km/(Km + 10Km) = 10/11 = 0.91
Chap. 6. Problem 12. The Vmax and Km for prostaglandin endoperoxide synthase, and the type of inhibition that occurs in the presence of ibuprofen, can easily be determined from double-reciprocal plots of the kinetic data. To construct these graphs, we first have to calculate 1/[S] and 1/V0 from the data in the table (next slide).
Chap. 6. Problem 12 (cont.). Part (a). The double-reciprocal plots obtained with the transformed kinetic data are The y-intercept is equivalent to 1/Vmax; the x-intercept is equivalent to -1/Km. In the absence of inhibitor, Vmax and Km therefore are: 1/Vmax = 0.0190 min/mM and Vmax = 52.6 mM/min -1/Km = -1.7 mM-1 and Km = 0.59 mM
Chap. 6. Problem 12 (cont.). Part (b). The plot obtained with ibuprofen intersects the y-axis at the same value of 1/Vmax as obtained without the inhibitor. Thus the Vmax for the reaction is unchanged. The inhibitor plot intersects the x-axis at a smaller value of -1/Km (that is a greater value of Km). The plot in the presence of the inhibitor therefore reveals that ibuprofen is a competitive inhibitor of prostaglandin endoperoxide synthase. (See Fig. 1 in Box 6-2 of the textbook).
Chap. 6. Problem 15. The turnover number for an enzyme is the number of substrate molecules converted to product per unit time by a single enzyme molecule when the enzyme is saturated with substrate. The turnover number, kcat is calculated from the relationship kcat = Vmax/Et, where Et is the total moles of enzyme. The turnover number in units of min-1 can be obtained if we first convert the weights of the enzyme and substrate to molar amounts: Vmax = (0.30 g/min)/(44 g/mol) = 6.8 x 10-3 mol/min Et = (10 g)(1 g/106g)/30,000 g/mol = 3.3 x 10-10 mol The turnover number now can be obtained by dividing the moles of CO2/min by the moles of enzyme present, or Kcat = (6.8 x 10-3 mol/min)/(3.3 x 10-10 mol) = 2.0 x 107 min-1
Chap. 6. Problem 18. To determine acid phosphatase activity in the blood that is derived from the prostate gland, one would need to measure activity in the presence and absence of tartrate. If for example 100 U of activity were measured in the absence of tartrate, and 10 U were measured in its presence, then the blood would contain 90 U of activity originating from the prostate. In other words, the difference between the two activities represents the activity of acid phosphatase originating from the prostate gland.
Chap. 6. Problem 21. The pH optimum of lysozyme (pH 5.2) is approximately 1 unit below and above the respective pKas of Glu35 and Asp52 in the active site. Thus, about 90% of the Glu35 side-chain would be present in its conjugate acid (protonated) form, and 90% of the Asp52 side-chain would be present in its conjugate base (ionized) form at the pH optimum. Because activity declines above pH 5.2, it suggests that the Glu35 side-chain must be in its protonated form for catalysis. Likewise, because activity declines below pH 5.2, Asp52 must be in its ionized form for catalysis.