Download

1 / 122
1.23k likes | 1.32k Views
Explore the inheritance of autosomal dominant disorders, including examples like familial hypercholesterolemia and achondroplasia. Learn about new mutations, fitness implications, and sex-limited phenotypes in these genetic conditions.

E N D
Patterns of Single-Gene Inheritance Lecture 3
Autosomal Dominant Inheritance • More than half of all mendelian disorders are inherited as AD traits. • The incidence of some autosomal dominant disorders is high, e.g., familial hypercholesterolemia,myotonic dystrophy, Huntington disease, neurofibromatosis, and polycystic kidney disease.
AD disorders are individually much less common, in aggregate their total incidence is appreciable. • The burden of autosomal dominant disorders is increased because of their hereditary nature; they become problems for whole kindreds, often through many generations. • In some cases, the burden is compounded by social difficulties resulting from physical or mental disability.
The risk and severity of dominantly inherited disease in the offspring depend on whether one or both parents are affected and whether the trait is strictly dominant or incompletely dominant. • Denoting D as the mutant allele and d as the normal allele, matings that produce children with an autosomal dominant disease can be between two heterozygotes (D/d) for the mutation or, more frequently, between a heterozygote for the mutation (D/d) and a homozygote for a normal allele (d/d):
Offspring of D/d x d/d are approximately 50% D/d and 50% d/d. • Each pregnancy is an independent event, not governed by the outcome of previous pregnancies. • Thus, within a family, the distribution of affected and unaffected children may be quite different from the theoretical expected ratio of 1:1, especially if the sibship is small.
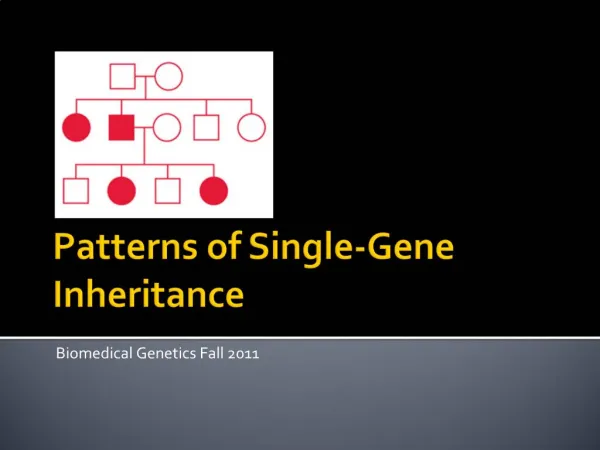
A pedigree showing typical inheritance of a form of progressive sensorineural deafness (DFNA1) inherited as an autosomal dominant trait.
Achondroplasia, an AD disorder that often occurs as a new mutation. • Note small stature with short limbs, large head, low nasal bridge, prominent forehead, and lumbar lordosis in this typical presentation.
In medical practice, homozygotes for dominant phenotypes are not often seen because matings that could produce homozygous offspring are rare. • Which mating can produce a D/D homozygote? • Practically, only the mating of two heterozygotes need be considered because D/D homozygotes are very rare and generally too severely affected to reproduce (fitness =0).
Incompletely Dominant Inheritance • Achondroplasia: incompletely dominant skeletal disorder of short-limbed dwarfism and large head. • Most achondroplastics have normal intelligence and lead normal lives within their physical capabilities. • A homozygous child of two heterozygotes is often recognizable on clinical grounds alone; much more severely affected and commonly do not survive the immediate postnatal period.
A pedigree of a mating between two individuals heterozygous for the mutation that causes achondroplasia. The deceased child, individual III-3, was a homozygote and died soon after birth.
Another example is AD familial hypercholesterolemia, leading to premature coronary heart disease. • The rare homozygotes have a more severe disease, with an earlier age at onset and much shorter life expectancy.
Cutaneous xanthomas in a familial hypercholesterolemia homozygote.
New Mutation in Autosomal Dominant Inheritance • In typical AD inheritance, every affected person in a pedigree has an affected parent, who also has an affected parent, and so on as far back as the disorder can be traced or until the occurrence of an original mutation. • This is also true, for X-linked dominant pedigrees. In fact, most dominant conditions of any medical importance come about not only through transmission of the mutant allele but also through inheritance of a spontaneous, new mutation in a gamete.
Relationship Between New Mutation and Fitness in Autosomal Dominant Disorders • Once a new mutation has arisen, its survival in the population depends on the fitness of persons carrying it. • There is an inverse relation between the fitness of a given AD disorder and the new mutation. • At one extreme are disorders that have a fitness of zero, and the disorder is referred to as a genetic lethal. Must be due to new mutations.
The affected individual will appear as an isolated case in the pedigree. • If the fitness is normal, the disorder is rarely the result of fresh mutation; and the pedigree is likely to show multiple affected individuals.
Sex-Limited Phenotype in Autosomal Dominant Disease • AD phenotypes may also demonstrate a sex ratio that differs from 1:1. • Extreme divergence of the sex ratio is seen in sex-limited phenotypes, in which the defect is autosomally transmitted but expressed in only one sex. • An example is male-limited precocious puberty (familial testotoxicosis), an AD disorder in which affected boys develop secondary sexual characteristics and undergo an adolescent growth spurt at about 4 years of age.
In some families, the defect is in the gene that encodes the receptor for luteinizing hormone (LCGR); these mutations constitutively activate the receptor's signaling action even in the absence of its hormone. • The defect is not manifested in heterozygous females. • Although the disease can be transmitted by unaffected females, it can also be transmitted directly from father to son, showing that it is autosomal, not X-linked.
Males with precocious puberty due to activating LCGR mutations have normal fertility, and numerous multigeneration pedigrees are known. • For disorders in which affected males do not reproduce, however, it is not always easy to distinguish sex-limited autosomal inheritance from X-linkage because the critical evidence, absence of male-to-male transmission, cannot be provided. • In that case, other lines of evidence, especially gene mapping to learn whether the responsible gene maps to the X chromosome or to an autosome, can determine the pattern of inheritance and the consequent recurrence risk.
Pedigree pattern (part of a much larger pedigree) of male-limited precocious puberty in the family of the child shown in Figure 7-14. This autosomal dominant disorder can be transmitted by affected males or by unaffected carrier females. Male-to-male transmission shows that the inheritance is autosomal, not X-linked. Because the trait is transmitted through unaffected carrier females, it cannot be Y-linked.
Characteristics of Autosomal Dominant Inheritance • The phenotype usually appears in every generation, each affected person having an affected parent. • Exceptions or apparent exceptions (1) cases originating from fresh mutations and (2) cases in which the disorder is not expressed (nonpenetrant) or is expressed only subtly in a person who has inherited the responsible mutant allele. • Any child of an affected parent has a 50% risk of inheriting the trait. • This is true for most families, in which the other parent is phenotypically normal. Wide deviation from the expected 1:1 ratio may occur by chance in a single family.
Phenotypically normal family members do not transmit the phenotype to their children. • exceptions. • Males and females are equally likely to transmit the phenotype, to children of either sex. In particular, male-to-male transmission can occur, and males can have unaffected daughters. • A significant proportion of isolated cases are due to new mutation. The less the fitness, the greater is the proportion due to new mutation.
X-LINKED INHERITANCE • Phenotypes determined by genes on the X have a characteristic sex distribution and a pattern of inheritance that is usually easy to identify. • Approximately 1100 genes are thought to be located on the X chromosome, of which approximately 40% are presently known to be associated with disease phenotypes
There are only two possible genotypes in males and three in females with respect to a mutant allele at an X-linked locus. • A male with a mutant allele at an X-linked locus is hemizygous for that allele, whereas females may be homozygous for either the wild-type or mutant allele or may be heterozygous. • For example, if XH is the wild-type allele for the gene for coagulation factor VIII and a mutant allele, Xh, causes hemophilia A, the genotypes expected in males and females would be as follows:
X Inactivation, Dosage Compensation, and the Expression of X-Linked Genes • The clinical relevance of X inactivation is profound. It leads to females having two cell populations, one in which one of the X chromosomes is active, the other in which the other X chromosome is active. • For example, in Duchenne muscular dystrophy, female carriers exhibit typical mosaic expression, allowing carriers to be identified by dystrophin immunostaining. • Depending on the pattern of random X inactivation of the two X chromosomes, two female heterozygotes for an X-linked disease may have very different clinical presentations because they differ in the proportion of cells that have the mutant allele on the active X in a relevant tissue (as seen in manifesting heterozygotes).
Immunostaining for dystrophin in muscle specimens. A, A normal female (magnification ×480). B, A male with Duchenne muscular dystrophy (×480). C, A carrier female (×240). Staining creates the bright lines seen here encircling individual muscle fibers. Muscle from DMD patients lacks dystrophin staining. Muscle from DMD carriers exhibits both positive and negative patches of dystrophin immunostaining, reflecting X inactivation
Recessive and Dominant Inheritance of X-Linked Disorders • X-linked "dominant" and "recessive" patterns of inheritance are distinguished on the basis of the phenotype in heterozygous females. Some X-linked phenotypes are consistently expressed in carriers (dominant), whereas others usually are not (recessive). • The difficulty in classifying an X-linked disorder as dominant or recessive arises because females who are heterozygous for the same mutant allele in the same family may or may not demonstrate the disease, depending on the pattern of random X inactivation and the proportion of the cells in pertinent tissues that have the mutant allele on the active versus inactive chromosome.
X-Linked Recessive Inheritance • The inheritance of X-linked recessive phenotypes follows a well-defined and easily recognized pattern. • An X-linked recessive mutation is typically expressed phenotypically in all males who receive it but only in those females who are homozygous for the mutation. • X-linked recessive disorders are generally restricted to males and rarely seen among females (except for manifesting heterozygotes).
Hemophilia A is a classic X-linked recessive disorder in which the blood fails to clot normally because of a deficiency of factor VIII. • The hereditary nature of hemophilia and even its pattern of transmission have been recognized since ancient times, and the condition became known as the "royal hemophilia" because of its occurrence among descendants of Britain's Queen Victoria, who was a carrier. • If a hemophiliac mates with a normal female ? • Now assume that a daughter of the affected male mates with an unaffected male ?
Pedigree pattern demonstrating an X-linked recessive disorder such as hemophilia A, transmitted from an affected male through females to an affected grandson and great-grandson.
Homozygous Affected Females • Relevant for X-linked color-blindness, a relatively common X-linked disorder (an affected male x a carrier female). • Most X-linked diseases are so rare, unusual for a female to be homozygous unless parents are consanguineous • Affected male x carrier female
Homozygous affected female Consanguinity in an X-linked recessive pedigree for red-green color blindness, resulting in a homozygous affected female
Manifesting Heterozygotes and Unbalanced Inactivation for X-linked Disease • Rare, a female carrier of a recessive X-linked allele has phenotypic expression of disease = manifesting heterozygote. • Have been described for many X-linked recessive disorders, e.g., color-blindness, hemophilia A & B, DMD, Wiskott-Aldrich syndrome (an X-linked immunodeficiency), etc. • Whether a female heterozygote will be a manifesting heterozygote depends on a number of factors:
First, the fraction of cells in which the normal/mutant allele happens to remain active (unbalanced or skewed X-inactivation). • Second, depending on the disorder in question, females can have very different degrees of disease penetrance and expression, even if their degree of skewed inactivation is the same, because of underlying physiological functioning of genes e.g., • In Hunter syndrome (iduronate sulfatase deficiency), cells with normal allele on active X can export enzyme to extracellular space, picked up by cells in which mutant allele on active X and defect is corrected in those cells
So, penetrance for Hunter syndrome in heterozygous females is extremely low even when X-inactivation deviates from expected random 50%:50% pattern • On the other hand, nearly half of all female heterozygotes for fragile-X syndrome show developmental abnormalities. • In addition to manifesting heterozygotes, the opposite pattern of skewed inactivation can also occur.
Characteristics of X-Linked Recessive Inheritance • The incidence of the trait is much higher in males. • Heterozygous females are usually unaffected, exception? • The gene responsible is transmitted from an affected man through all his daughters. Any of his daughters' sons has a 50% chance of inheriting it. • The mutant allele is ordinarily never transmitted directly from father to son. • The mutant allele may be transmitted through a series of carrier females; if so, the affected males in a kindred are related through females. • A significant proportion of isolated cases are due to new mutation.
X-linked Dominant Inheritance • Regularly expressed in heterozygotes • No male-to-male transmission • For a fully penetrant XD pedigree, all daughters and none of sons of affected males are affected. • Pattern of inheritance through female is no different from AD. • The expression is usually milder in females, who are almost always heterozygotes. Thus, most XD disorders are incompletely dominant. • Only a few genetic disorders are classified as XD.
E.g., X-linked hypophosphatemic rickets (a.k.a. vitamin D-resistant rickets) • Defective gene product is one of the endopeptidases that activate or degrade a variety of peptide hormones • Both sexes are affected but, serum phosphate level is less depressed and rickets less severe in heterozgous females.
Pedigree pattern demonstrating X-linked dominant inheritance
X-linked Dominant Disorders with Male Lethality • Some rare genetic defects expressed exclusively or almost exclusively in females appear to be XD lethal in males before birth • Typical pedigrees: transmission by affected female affected daughters, normal daughters, normal sons in equal proportions (1:1:1) • Rett syndrome meets criteria for an XD that is usually lethal in hemizygous males. The syndrome is characterized by normal prenatal and neonatal growth and development, followed by rapid onset of neurological symptoms and loss of milestones between 6 and 18 months of age.
Rett syndrome cont. • Children become spastic and ataxic, develop autistic features and irritable behavior with outbursts of crying, and demonstrate characteristic purposeless wringing or flapping movements of hands and arms. • Head growth slows and microcephaly develops. Seizures are common (~50%) • Surprisingly, mental deterioration stops after a few years and the patients can then survive for many decades with a stable but severe neurological disability. • Most cases caused by spontaneous mutations in an X-linked MECP2 gene encoding methyl CpG binding protein 2. ? Thought to reflect abnormalities in regulation of genes in developing brain.
Typical appearance and hand posture of girls with Rett syndrome
Rett syndrome cont. • Males who survive with the syndrome usually have two X chromosomes (as in 47,XXY or in a 46,X,der(X) male with the male determining SRY gene translocated to an X) or are mosaic for a mutation that is absent in most of their cells • There are a few apparently unaffected women who have given birth to more than one child with Rett syndrome. ? X-inactivation pattern in a heterozygous female. ? Germline mosaic ?
Pedigree pattern demonstrating an X-linked dominant disorder, lethal in males during the prenatal period.
Characteristics of X-Linked Dominant Inheritance • Affected males with normal mates have no affected sons and no normal daughters. • Both male and female offspring of a heterozygous female have a 50% risk of inheriting the phenotype. The pedigree pattern is similar to that seen with autosomal dominant inheritance. • Affected females are about twice as common as affected males, but affected females typically have milder (although variable) expression of the phenotype.
New Mutation in X-Linked Disorders • In males, genes for X-linked disorders are exposed to selection that is complete for some disorders, partial for others, and absent for still others, depending on the fitness of the genotype. • Patients with hemophilia have only about 70% as many offspring as unaffected males do; that is, the fitness of affected males is about 0.70. • Selection against mutant alleles is more dramatic for X-linked disorders such as DMD. DMD is currently a genetic lethal because affected males usually fail to reproduce. It may, of course, be transmitted by carrier females, who themselves rarely show any clinical manifestation of the disease. • New mutations constitute a significant fraction of isolated cases of many X-linked diseases. When patients are affected with a severe X-linked recessive disease, such as DMD, they cannot reproduce (i.e., selection is complete), and therefore the mutant alleles they carry are lost from the population. Because the incidence of DMD is not changing, mutant alleles lost through failure of the affected males to reproduce are continually replaced by new mutations.
PSEUDOAUTOSOMAL INHERITANCE • Pseudoautosomal inheritance describes the inheritance pattern seen with genes in the pseudoautosomal region. • Alleles for genes in the pseudoautosomal region can show male-to-male transmission, and therefore mimic autosomal inheritance, because they can cross over from the X to the Y during male gametogenesis and be passed on from a father to his male offspring.
Dyschondrosteosis, a dominantly inherited skeletal dysplasia with disproportionate short stature and deformity of the forearm, is an example of a pseudoautosomal condition inherited in a dominant manner. • A greater prevalence of the disease was seen in females as compared with males, suggesting an X-linked dominant disorder, but the presence of male-to-male transmission clearly ruled out strict X-linked inheritance. • Mutations in the SHOX gene encoding a homeodomain-containing transcription factor have been found responsible for this condition. • SHOX is located in the pseudoautosomal region on Xp and Yp and escapes X inactivation.
Figure 7-22 Pedigree showing inheritance of dyschondrosteosis due to mutations in a pseudoautosomal gene on the X and Y chromosomes. The arrow shows a male who inherited the trait on his Y chromosome from his father. His father, however, inherited the trait on his X chromosome from his mother