Download

1 / 1
10 likes | 154 Views
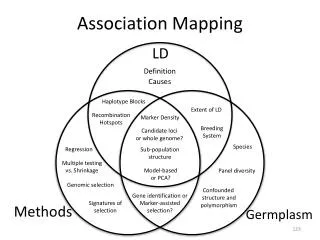
2. 4. 6. 8. 10. 12. 16. 18. 20. 22. 24. P lac. parS+/ *. yf p. parA1. parB. Mapping functional domains responsible for DNA association and dynamic movement in ParA1 of Streptomyces coelicolor.

E N D
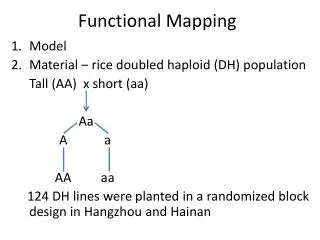
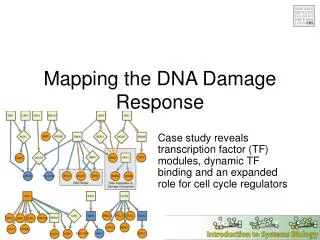
2 4 6 8 10 12 16 18 20 22 24 Plac parS+/ * yfp parA1 parB Mapping functional domains responsible for DNA association and dynamic movement in ParA1 of Streptomyces coelicolor Kuan-Yi Wu1 (巫冠毅), Yuan-Chi Lin2 (林婉琦), Jui-Heng Tseng2 (曾睿恆), Ting-Sung Hsieh1 (謝廷松) and Yu-Ling Shih1, 2 (史有伶) 1Institute of Biochemical Science, National Taiwan University, Taipei2Institute of Biological Chemistry, Academia Sinica, Taipei Introduction The Par (partition) system is known to play critical roles in stable inheritance of low-copy number plasmids in bacteria. In addition, it is one of the mechanism contributing to chromosome segregation during bacterial cell division. The Par system is comprised of two protein components, ParA and ParB, and a centromere-like parS DNAsite. ParA belongs to the MinD/ParA subgroup of the Walker-type ATPase family that is typified by their cytoskeleton properties, such as dynamic movement in the cell and formation of protein filaments in vitro. ParB was suggested to mediate plasmid pairing during DNA partition through specific binding to the parS sites. Specific binding of ParA to DNA is achieved via its interaction with the ParB-parS complex. In Streptomyces coelicolor, chromosome segregation in aerial hypha during sporulation is mediated by the Par system. However, the cytoskeleton properties of ParA1 and the mechanism to drive chromosome partitionhave not been confirmed. In this study, we aim at addressing these questions by examining (1) behavior of ParA1 in a heterologous host E. coli; (2) effects of ParA1 mutants on its dynamic behavior; (3) ability of the Scoe Par system in maintaining plasmid stability. ParA1domain organization and associating functions (A) N-terminus (A) (C) YFP DAPI Merge DIC K39E G40V K39E K44E 180º R218 D68A R247 (B) Results D154A N-terminal domain of ParA1 is required for nucleoid association in E. coli (D) Live cell Fixed cell (C) (A) DIC YFP DAPI Merge YFP DAPI Merge DIC R8E R19E Yfp::ParA1 R26E (B) (D) DIC YFP DAPI merge YFP DIC Merge Yfp::ParA1NC R19E+R26E Figure 5. Background information of ParA1 and nucleoid localization of the mutant ParA1 proteins. (A) Protein structure of a ParA homolog Soj from Thermus thermophilus (PDB entry 2BEJ). Blue: Walker A box; Orange: Box IV; Green: Walker B box; Yellow: central helix. (B) Summary of domain functions delivered by specific residues in ParA1. (C) Mutations in the Walker A (K39, G40, K44) and Walker B (D154) boxes caused loss of nucleoid localization in E. coli, but mutation D68 in the Box IV had no effect. (D) Only R31E mutation in the N-terminal domain of ParA1 failed nucleoid localization. (E) Mutations (R218, R247) in the C-terminal domain abolished nucleoid targeting ability of ParA1. Q29E Figure 1.Localization of full-length ParA1(A, C) and N-terminal (amino acids 1-32) truncated ParA1NC (B, D) in E. coli. The experiments were done by tagging the yellow fluorescent protein (Yfp) to ParA1 or ParA1NC. The cellular localization of these constructs were studied in wild-type E. coli cells. Nucleoids were stained with DAPI. Note the filamentous structures (arrows) wrapping around the nucleoid (C) or the cell (D) were found in fixed samples. R31E (E) R218E Stochastic movement of ParA1 and ParA1NC in live cells R247E (A) Yfp::ParA1 3.2 5.6 8 8.4 7.8 4 7.4 6.4 10 10.2 10.4 Sec Mutations in ParA1 affect intermolecular interactions (B) Yfp::ParA1NC (A) (B) pT25 cya’’ parA1 pT18 parA1 cya’ 0 Sec Figure 6. Interactions between mutant ParA1 are monitored by bacterial two-hybrid system. (A) Interaction between mutant ParA1 proteins in the bacterial two hybrid system is expressed in Millerunits (n≥3). (B) Illustration of constructs used in the experiment. Figure 2. Dynamic movement of ParA1 (A) andParA1NC (B). The results indicate that dynamic movement of the YFP taggedParA1NCis independent of nucleoid targeting. Asterisks indicate positions of the most concentrated fluorescence in cells. Miller unit ParB and parS are required for localizing ParA1into discrete foci pT25-parA WT NC WT R31EK39E G40V K44E D68A D154A R218E R247E ParBParB pT18-parA WT NC NC R31EK39E G40V K44ED68A D154A R218E R247E WTD68A (B) (A) Curing of miniF (sopABC-) plasmid by parA1BS of S. coelicolor YFP sopA sopB (A) (B) sopC YFP miniF plasmid DAPI DAPI parB cat parA1 Merge parS Merge Recombinant miniF 12 3 (C) DIC DIC Figure 3.Formation of discrete Yfp-ParA1 foci in E. coli requires functional ParB and parS.(A) In the presence of ParB and parS, ParA1localized into discrete foci (arrows) as well as targeting to the nucleoids. (B) The focal localization was diminished when the parS site was disrupted by introducing point mutation into the parS site but ParB remained unchanged, suggesting formation of the ParA1 foci is caused by nucleation of ParA1 on the ParB-parS complex formed on the plasmid. Figure 7. (A) Effects of substituting sopABC of the miniF plasmid with parA1BS of S. coelicolor on maintaining plasmid stability.(B) Scheme of substituting the sop system on miniF via the RED mediated homologous recombination. (C) Illustration of plasmid loss over generation due to partition defects. Helical structure formation ofParA1NC is independent of the Min system and the actin homolog MreB Summary (B) (A) ΔminCDE/Plac- yfp::parA1NC time image YFP DIC Merge • The ability of helical structure formation and dynamic movement of ParA1can be separated from nucleoid association. The result supports the idea that the cytoskeletal property of the MinD/ParA group of proteins is independent of their biological functions. • ParA1 can be dissected into three functional domains, including the N- and C-terminal domains and the ATPase domain. • While the N- and C-terminal domains of ParA1 are required for nucleoid targeting, the ATPase domain participates in interactions between ParA1 and ParA1, ParA1 and ParB, as well as ParA1 and nucleooids. • parA1BS of S. coelicolor can substitute the endogenous sopABC on the low-copy number miniF plamid to maintain stable inheritance of the plasmid. Figure 4. Formation of the long-range helical filaments of ParA1NC in the absence of the Min system and MreB. (A) The Yfp-ParA1NC(arrow) was expressed in a minCDE knockout strain of E. coli. (B)The Yfp-ParA1NCwas expressed in a wild-type E. coli strain treated with a compound A22, S-(3,4 dichlorobenzyl) isothiourea, to disrupt the helical structure formation of MreB. A22 acts through blocking the ATP-binding pocket in MreB. Acknowledgement We thank useful discussions and helps from Dr. Carton Chen’s laboratory. This work is supported by Academia Sinica (AS97-FP-M02-2, AS98-CDA-L07).