Download

1 / 30
310 likes | 469 Views
The dynamics of MHC class I antigen presentation. Neil Dalchau Computational Science Laboratory Biological Computation Group. Computational Life and Medical Sciences Network Second Annual Symposium 28 th June 2012, London. Immune System Computation.

E N D
The dynamics of MHC class I antigen presentation Neil Dalchau Computational Science Laboratory Biological Computation Group Computational Life and Medical Sciences Network Second Annual Symposium 28th June 2012, London
Immune System Computation • The immune system solves a multi-scale problem • Perceives intracellular protein content • Translates this into a co-ordinated multi-cellular response • The response is specific to the invader • Decision-making is required to determine which cells to kill, and which antibodies to produce
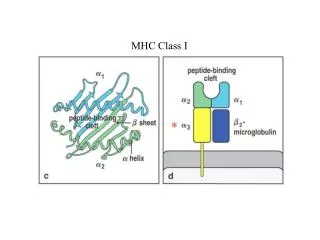
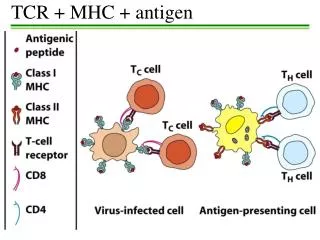
An insanely variable interaction MPLSYQHFRKLLLLDDEAGPLEEELPRLA.... • T cell activation relies on a productive interaction between its receptor and a peptide-MHC complex • MHC class I binds and presents peptides of length 8 – 13 • Each T cell has a unique TCR, that sees (at least some of) the peptide • Therefore, an enormous diversity of T cells is required to cover the variability between peptides …AND distinguish between self and non-self Protein sequence MHC class I molecule MHC class I molecule S__N_E_L SIINFEKL Protein fragment (peptide) TCR TCR
(Within-host) Immunodominance • Despite the large number of possible interactions, the immune response is dominated by a small set of peptides • Difficult to explain without some quantitative thinking • How does biology compute optimal responses over combinatorially diverse interactions? Source: Ian York’s blog. Adapted from Kotturiet al., J Immunology 2008
A computational model of immunodominance? • Seeking to develop a multi-scale model of the CTL response • Modular approach looking at the multiple factors involved • Several associated with cell surface peptide-MHC • Several associated with T cell populations • Challenges: • Heterogeneous data (human vs. mouse vs. macaque, in vivo, in vitro, etc.) • Incorporating experimental data (Bayesian inference) • Scalable modelling methodologies (domain-specific languages)
MHC class I: Kinetics, structure and function • How do MHC class I molecules select epitopes to present to T cells? • How do chaperone molecules (i.e. Tapasin) aide the selection of an “optimal” ligand? • How do genetic polymorphisms in the MHC locus affect interactions with chaperones, and thus influence the ability to present antigen? Denise Boulanger Rachel Carter Andy van Hateren Tim Elliott Joern Werner Alistair Bailey
Cell surface abundance depends on a number of factors: • peptide-MHC unbinding rate • effects of tapasin • peptide abundance
Peptide filtering (If )
Peptide filtering Tapasin unbinding (If ) Enhanced peptide dissociation • Tapasineither • acceleratespeptide unbinding • decelerates cell surface transit (ER retention)
Peptide filter relation • Tapasin confers an additional filter, enhancing presentation proportional to affinity • In steady state, cell surface abundance of a specific peptide-MHC complex can be approximated by a single equation
Peptide optimisation over time • Tapasin helps MHC class I form stable complexes with peptides • Different MHC alleles vary in their tapasin-dependency
Model selection and parameterisation • Can we use the model to learn how HLA-B molecules vary? • What is is allele-specific | Data? • Bayes’ Theorem • Bayesian Information Criterion • Parameter estimation • Adaptive Markov Chain Monte Carlo (MCMC)sampler BIC = f1(deviation from data) + f2(no. of parameters)
Model selection and parameterisation • Can we use the model to learn how HLA-B molecules vary? • What is is allele-specific | Data?
Peptide optimisation over time • Tapasin helps MHC class I form stable complexes with peptides • Different MHC alleles vary in their tapasin-dependency
Relating structure to understanding • Conformational change is fundamental to how many receptor proteins work • Haemoglobin • MHC class I • Hypothesis: Tapasin achieves improved peptide optimization by altering the conformation of the MHC molecule • Our MHC class I model only implicitly incorporates structural variability • How can we explicitly interpret conformational change of MHC class I molecules?
Sequence ⇒ Structure ⇒ Function GSHSMRYFYTAMSRPGRGEPRFITVGYVDDTLFVRFDSDATSPRKEPRAPWIEQEGPEYWDRETQISKTNTQTYRENLRTALRYYNQSEAGSHIIQRMYGCDVGPDGRLLRGYDQYAYDGKDYIALNEDLSSWTAADTAAQITQRKWEAARVAEQDRAYLEGLCVESLRRYLENGKETLQRADPPKTHVTHHPISDHEVTLRCWALGFYPAEITLTWQRDGEDQTQDTELVETRPAGDRTFQKWAAVVVPSGEEQRYTCHVQHEGLPKPLTLRWEP HLA-B*44:02 HLA-B*44:05 HLA-B*44:05 W147A
Sequence ⇒ Structure ⇒ Function GSHSMRYFYTAMSRPGRGEPRFITVGYVDDTLFVRFDSDATSPRKEPRAPWIEQEGPEYWDRETQISKTNTQTYRENLRTALRYYNQSEAGSHIIQRMYGCDVGPDGRLLRGYDQYAYDGKDYIALNEDLSSWTAADTAAQITQRKWEAARVAEQDRAYLEGLCVESLRRYLENGKETLQRADPPKTHVTHHPISDHEVTLRCWALGFYPAEITLTWQRDGEDQTQDTELVETRPAGDRTFQKWAAVVVPSGEEQRYTCHVQHEGLPKPLTLRWEP HLA-B*44:02 HLA-B*44:05 HLA-B*44:05 W147A
A structural intermediate with Alistair Bailey, Joern Werner and Tim Elliott (Southampton) Hypothesis: Tapasin achieves improved peptide optimization by altering the conformation of the MHC molecule Basic Model Mathematically equivalent Flytrap Model “Open” “Closed”
A structural intermediate • What is: • Pr(Basic | data)? • Pr(Flytrap | data)? • Pr(Inverse Flytrap | data)? • Bayesian Inference (Filzbach) • Can we use model predictions to design experiments that distinguish between our hypotheses? Rachel Carter
Upstream of MHC binding Protein TAP Proteolysis TAP Binding TAP Peptides QIOSJKLSFVINMC...PLLLLXWFJKLSFLK…SKLAALALLLSNAA… JKLSFVIN WFJKLSFL ER
Bringing dynamics to static models • Many algorithms for predicting peptide-MHC affinity …proteasomal degradation… …and TAP binding • We can take these predictions and convert their predictions into rates (ball-park estimates)
Dynamic modelling of HIV recognition by the immune system • Model the lifecycle of the HIV virus (replication, cell entry/exit and protein abundance) • Analyse the proteasome in the context of viral protein processing and investigate the sufficiency of existing models • Create an integrated model of viral infection and MHC class I presentation on the surface of an antigen presenting cell to investigate virus spread • Investigate the down-regulation of peptide processing and filtering by viruses Peter Coveney
Outlook • Understanding immune system computation • Immunodominance • Sequence, structure, kinetics, function • Predictions for the community? • Could the model be useful for vaccine design? • Technological innovation • Parameter estimation (Filzbach) • Multi-scale modelling methodologies
Acknowledgements Tim Elliott Joern Werner Alistair Bailey • Neil Dalchau • Andrew Phillips • Stephen Emmott • Luca Cardelli
Biological Computation Group DNA Computing • Synthetic Biology • Development Immunology Biocharts SPiM DSD GEC Biological Modelling Engine