Download

1 / 21
210 likes | 315 Views
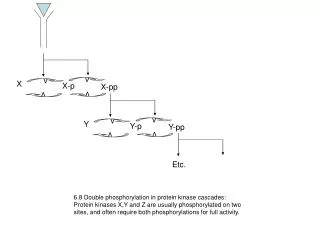
PROSITE contiene anche pattern ad ALTA OCCORRENZA, corti e aspecifici (modifiche post-traduzionali). Es. phosphorylation by CK2 [ST]-x(2)-[DE].

E N D
PROSITE contiene anche pattern ad ALTA OCCORRENZA, corti e aspecifici (modifiche post-traduzionali) Es. phosphorylation by CK2 [ST]-x(2)-[DE]
PROSITE: pattern funzionali – pattern relativi a corte seq con alta probabilità di accadimento (modificazioni e non funzioni) – profili – indicazioni su SPECIFICITA’ e SENSIBILITA’ del pattern (POTERE DIAGNOSTICo); • PRINTS: costruisce la signature di una famiglia codificata come fingerprint – si autoimplementa usando SwissProt partendo da un allineamento seme – punteggi basati sulla freq di vedere un aa in una posizione • BLOCKS:parte dalle famiglie di Prosite – punteggi dati con Blosum 62 – ogni blocco valutato sulla base di 2 punteggi: livello a cui 99,5% sono negativi e forza della ricerca (devono essere ben distinti e la forza maggiore del primo punteggio) • Pfam: fornisce info su domini strutturali codificate sottoforma di profili (metodo HMM)– ci sono poi indicazioni su repeats, sequenze segnale, regioni transmembrana, regioni a bassa complessità
perchè è interessante studiare la struttura di una proteina in molti casi è vero che solo l’analisi della struttura tridimensionale di una macromolecola può aiutarci a comprendere in quale modo e per quale motivo una determinata sequenza (avvolta in una specifica struttura) possa codificare una ben precisa funzione vediamo per esempio la struttura 3D della chimotripsina, mettendo in evidenza i residui della triade catalitica, che non sono contigui nella sequenza proteica la contiguità dei residui in struttura determina la funzione
Predizione della struttura di una proteina In generale, la sequenza amminoacidica di una proteina codifica la sua struttura tridimensionale dovrebbe quindi essere possibile disegnare un algoritmo per predire la struttura 3D di una proteina a partire dalla sua sequenza 1D questo è l’obiettivo più ambizioso e complesso della bioinformatica, e non è ancora stato raggiunto (reverse folding)
i metodi sperimentali per la determinazione della sequenza di una proteina sono estremamente rapidi (l’ordine di grandezza è il giorno) e relativamente economici la risoluzione della struttura tridimensionale di una proteina richiede invece l’uso di strumenti più complessi, e talvolta mesi di lavoro Swiss-Prot + TrEMBL circa 10 milioni di sequenze PDB (Protein Data Bank) circa 64.000 di strutture gran parte delle ricerche in biologia strutturale è quindi volta allo studio delle leggi fondamentali del folding delle proteine e la biologia computazionale dedica molte energie e risorse allo sviluppo di metodi per la predizione della struttura delle proteine
600000 500000 400000 300000 200000 100000 0 Sequenze Comparative Models Strutture Perchè predire la struttura terziaria? • In cifre: • 10 milioni sequenze proteiche • Piu‘ di 64000 strutture, di cui 6000 uniche • La distanza tra sequenze e strutture note si sta allargando. • Metodi computazionali • Veloci ( poche ore/giorni), poco costosi (PC) • Soluzioni corrette ca. nel 60% dei casi. • Risoluzione più bassa, ma spesso sufficiente per spiegare la funzione e dettagli strutturali delle proteine • Osservazione: La sequenza si evolve più rapidamente della struttura (Chothia & Lesk, 1986) • Numero limitato di fold (< 1,000? )
perchè è interessante studiare la struttura di una proteina il confronto tra sequenze proteiche può essere utilizzato per mettere in luce relazioni evolutive tra proteine e la similarità tra sequenze può essere utilizzata come una misura della distanza evolutiva tra gli organismi come abbiamo visto, le proteine che si confrontano possono talvolta essere così diverse che diventa difficile metterne in evidenza la comune origine evolutiva attraverso il solo confronto tra sequenze cambiamenti nella struttura delle proteine sono invece più conservativi: l’evoluzione delle proteine avviene in modi che in genere non alterano il ripiegamento (fold) della struttura proteica, che quindi può conservare tracce di un’origine comune
Come si può studiare la struttura di una proteina i metodi sperimentali classici per la risoluzione della struttura tridimensionale di una proteina sono: • la cristallografia a raggi X • la spettroscopia a risonanza magnetica e nucleare (Nuclear Magnetic Resonance, NMR)
diffrazione ai raggi X cellula batterica DNA esogeno NMR plasmide formazione di cristalli moltiplicazione del clone purificazione della proteina
Cristallografia a raggi X La proteina, cristallizzata, viene bombardata con un raggio di fotoni collimati ad alta energia. I fotoni vengono diffratti in modo differente a seconda del tipo di atomo che colpiscono. I raggi diffratti vengono raccolti formando un quadro(pattern) di diffrazione Il pattern di diffrazione viene usato per ricostruire le coordinate dei singoli atomi che compongono la macromolecola e quindi la sua struttura 3D. I raggi X interagiscono quasi esclusivamente con gli elettroni presenti nella materia e non con i nuclei. Una struttura ai raggi X è quindi un’immagine della densità elettronica dell’oggetto in analisi
Cristallografia a raggi X Risoluzione ottenibile su piccole molecole organiche 1Å. In generale proteine cristallizate hanno grado di organizzazione più basso che limitano la risoluzione (2-3.5Å). Questo è dovuto anche all’alta idratazione dei cristalli (40-60% di acqua). Una tale risoluzione non è sufficiente per rivelare la posizione dei singoli atomi, ma è sufficiente per tracciare l’andamento e la disposizione dello scheletro covalente della proteina. Occorre quindi conoscere la sequenza primaria in modo da adattare la mappa della densità elettronica alla sequanza aminoacidica. • Limiti del metodo: • Cristallizzare proteine e` difficile • Ricavare la struttura dal pattern di diffrazione e` computazionalmente complesso • L’informazione che si ottiene e` statica, mentre la conforomazione di una proteina in soluzione varia nel tempo • B factor (fattore di temperatura): indice “indiretto” di flessibilita’ strutturale
In un cristallo di proteine, in ogni cella (sito reticolare) si trova una proteina = somma di densità elettroniche dovute ai singoli atomi
Il risultato sperimentale diretto di un’ analisi cristallografica è una mappa di densità elettronica e non il modello atomico che voi state osservando!!! Se vi sono errori nelle strutture cristallografiche questi sono dovuti all’interpretazione (soggettiva) delle mappe elettroniche da parte del cristallografo!!
PARAMETRI X VALUTAZIONE DELL’INFORMAZIONE CONTENUTA NELLA STRUTTURA X-RAY: Risoluzione (R) della mappa di densità elettronica (tra 3 e 1.5 Å). Strutture con R > 2.5 Å dettagli solo sul backbone, R < 2 Å dettagli su atomi catene laterali R-value e R-free: parametri associati al livello di “discordanza” tra modello atomico e mappa di densita’ elettronica (devono essere R-value < 20% e R-free < 30%). Forniscono, in ultima istanza, indicazioni su quanto differiscono tra loro mappa di densita’ elettronica osservata (sperimentale) e mappa di densita’ elettronica calcolata dal modello della struttura finale.
Spettroscopia a risonanza magnetica nucleare(NMR) Permette di ottenere informazioni sulla struttura di una molecola attraverso l’interazione con una radiazione elettromagnetica.Si basa sullo stesso principio della risonanza magnetica usata in medicina, usa onde radio. I nuclei atomici (elettricamente carichi) ruotano, con una velocità angolare quantizzata, creando un momento magnetico Immersi in un campo magnetico omogeneo esterno i momenti magnetici si allineano (“traballando” a causa del rumore termico). I nuceli vengono irradiati da onde radio (RF), l’effetto è di “disallineare” (tanto da farli “ribaltare”). È possibile rilevare quando i momenti magnetici dei vari nuclei (che continuano a ruotare) si inclinano completamente sul piano perpendicolare rispetto al campo magnetico applicatograzie ad un'antenna che capta le onde radio che questi generano ed è collocata perpendicolarmente al campo magnetico applicato.
Spettroscopia a risonanza magnetica nucleare(NMR) Ogni nucleo mostra le sue caratteristiche perchè ruota a velocità differente a seconda della sua posizione nella molecola e all'ambiente che gli atomi vicini gli fanno sentire e quindi risuona a frequenze radio diverse. Nuclei diversi risuonano a frequenze diverse. Ciò significa innanzitutto che un atomo di carbonio deve essere colpito da un'onda radio con frequenza diversa da quella necessaria ad un atomo di idrogeno per “ribaltarsi” di 90°, ma anche che atomi simili in ambienti diversi, come un atomo di idrogeno legato ad un atomo di ossigeno ed un atomo di idrogeno legato ad un atomo di carbonio si ribaltano a frequenze diverse. Questo è dovuto alla “schermatura” degli elettroni vicini • Caratteristiche: • studio delle proteine in soluzione (non occorre cristallizzarle) • alta risoluzione temporale (millisecondi) • informazioni sulle distanze interprotoniche non precise • la “proton signature” limita il metodo allanalsi di molecole “piccole” (<30 KD <250 residui) • informazioni su strutture di complessi proteici deducibili solo con il ricorso a metodi di modellistica guidati dai dati sperimentali
PDB: banca dati di strutture (Protein Data Bank) – ridondante – strutture ottenute sperimentalmente via X-ray o NMR Il file PDB http://www.pdb.org Esempio: Deossiemoglobina umana (1a3n) HEADER OXYGEN TRANSPORT 22-JAN-98 1A3N TITLE DEOXY HUMAN HEMOGLOBIN COMPND MOL_ID: 1; COMPND 2 MOLECULE: HEMOGLOBIN; COMPND 3 CHAIN: A, B, C, D; COMPND 4 BIOLOGICAL_UNIT: ALPHA-BETA-ALPHA-BETA TETRAMER SOURCE MOL_ID: 1; SOURCE 2 ORGANISM_SCIENTIFIC: HOMO SAPIENS; SOURCE 3 ORGANISM_COMMON: HUMAN; SOURCE 4 TISSUE: BLOOD; SOURCE 5 CELL: RED CELL KEYWDS OXYGEN TRANSPORT, HEME, RESPIRATORY PROTEIN, ERYTHROCYTE EXPDTA X-RAY DIFFRACTION AUTHOR J.TAME,B.VALLONE REVDAT 1 29-APR-98 1A3N 0 REMARK 1 REMARK 2 REMARK 2 RESOLUTION. 1.8 ANGSTROMS. REMARK 3 […]
tipo di atomo tipo di amminoacido coordinate X Y Z ATOM 1 N VAL A 1 10.720 19.523 6.163 1.00 21.36 N ATOM 2 CA VAL A 1 10.228 20.761 6.807 1.00 24.26 C ATOM 3 C VAL A 1 8.705 20.714 6.878 1.00 18.62 C ATOM 4 O VAL A 1 8.164 20.005 6.015 1.00 19.87 O ATOM 5 CB VAL A 1 10.602 22.000 5.966 1.00 27.19 C ATOM 6 CG1 VAL A 1 10.307 23.296 6.700 1.00 31.86 C ATOM 7 CG2 VAL A 1 12.065 21.951 5.544 1.00 31.74 C ATOM 8 N LEU A 2 8.091 21.453 7.775 1.00 16.19 N ATOM 9 CA LEU A 2 6.624 21.451 7.763 1.00 17.31 C ATOM 10 C LEU A 2 6.176 22.578 6.821 1.00 18.55 C ATOM 11 O LEU A 2 6.567 23.730 7.022 1.00 18.72 O ATOM 12 CB LEU A 2 6.020 21.707 9.129 1.00 18.34 C ATOM 13 CG LEU A 2 6.386 20.649 10.198 1.00 17.39 C ATOM 14 CD1 LEU A 2 5.998 21.119 11.577 1.00 17.99 C ATOM 15 CD2 LEU A 2 5.730 19.337 9.795 1.00 16.96 C ATOM 16 N SER A 3 5.380 22.237 5.852 1.00 15.02 N ATOM 17 CA SER A 3 4.831 23.237 4.928 1.00 16.59 C ATOM 18 C SER A 3 3.725 24.027 5.568 1.00 14.84 C ATOM 19 O SER A 3 3.095 23.717 6.591 1.00 14.40 O ATOM 20 CB SER A 3 4.308 22.429 3.727 1.00 16.47 C ATOM 21 OG SER A 3 3.076 21.786 3.991 1.00 14.91 O …