The Endocrine System
The Endocrine System. Chapter 17. Intercellular Communication. Cell. Endocrine. Target Cell. Blood. Hormone. Hormone. Neuroendocrine. Neuron. Target Cell. Blood. Hormone. Hormone. Paracrine. Cell. Interstitial Fluid. Target Cell. Hormone. Hormone. Autocrine. Cell.


The Endocrine System
E N D
Presentation Transcript
The Endocrine System Chapter 17
Intercellular Communication Cell Endocrine Target Cell Blood Hormone Hormone Neuroendocrine Neuron Target Cell Blood Hormone Hormone Paracrine Cell Interstitial Fluid Target Cell Hormone Hormone Autocrine Cell Interstitial Fluid Hormone
The endocrine system consists of the ductless glands. They are not connected anatomically. These glands secrete hormones into the blood. A hormone travels in the blood, signaling to distant target cells. Neurosecretory neurons release neurohormones. They are also distributed by the blood to target cells. Target cells are specific for each hormone or neurohormone. It is specific, as these cells have receptors that uniquely bind to a specific chemical messenger.
The endocrine and nervous systems are two regulatory systems of the body. The endocrine system mainly controls activities that require longer duration. This system has several overall functions. It regulates organic metabolism and water/electrolyte balance. It also induces adaptive changes to deal with stress. The endocrine system promotes smooth, sequential growth and development. Some hormones control reproduction while another one regulates red blood cell production. Some hormones regulate circulatory and digestive functions. A tropic hormone regulates the secretions of another endocrine gland. Tropic hormones are secreted by the anterior pituitary gland.
Classes of Hormones • Peptide & Protein Hormones • Steroid Hormones • Amine Hormones
Chemical Classification of Hormones Amine hormones are derived from tyrosine or tryptophan Include NE, Epi, thyroxine, melatonin Polypeptide/protein hormones are chains of amino acids Include ADH, GH, insulin, oxytocin, glucagon, ACTH, PTH Glycoproteins Long polypeptide bound to a carbohydrate group Include LH, FSH, TSH, hCG Steroids are lipids derived from cholesterol Include testosterone, estrogen, progesterone & cortisol 11-7
Peptide & Protein Hormones Gland/Tissue Hormones Hormones Gland/Tissue
Steroid Hormones Hormones Gland/Tissue
Amine Hormones Hormones Gland/Tissue
The mechanisms of hormone synthesis, storage, and secretion vary according to the class of hormone. Peptide hormones have precursors called preprohormones. They are made on ribosomes of the ER. In the Golgi complex they are converted to prohormones and, finally, active hormones. The Golgi complex concentrates these hormone into secretory vesicles. These hormones are released from endocrine cells by exocytosis. Cholesterol is the common precursor for all steroid hormones. A series of enzymatic steps modify this molecule into a different hormone in a specific endocrine cell. Only the precursor (cholesterol) is stored. The lipid-soluble hormone is not stored. The amine hormones are made from tyrosine.
All hormones are transported in the blood. However, they are not transported in the same way. Hydrophilic (water soluble) hormones are dissolved in the plasma. Most lipophilic (lipid soluble) hormones are bound reversibly to plasma proteins. These hormones are released by these proteins when they actively signal target cells. Hormones generally produce their effect by altering intracellular proteins. Hydrophilic hormones bind to receptors on the surface of target cells. Lipophilic hormones pass through target cell membranes and bind to receptors inside the target cell. A few hydrophilic hormones alter the permeability of the target cell’s membrane.
A hydrophilic hormone usually activates second-messenger systems. It binds to the target cell surface. This activates an intermediate G protein. This activates adenyl cyclase which converts intracellular ATP to cyclic AMP. Cyclic AMP triggers steps that alter the activity of a protein which is often an enzyme. This produces a physiological response in the target cell. A lipophilic hormone stimulates a gene, promoting protein synthesis. This kind of hormone passes through the target cell membrane. It binds with a receptor that binds to DNA. This turns on a gene. This gene makes RNA which makes a specific protein at the ribosome. This hormone changes the physiological response in the target cell. Compared to neural activity, the action of either class of hormone is usually slow and prolonged. Hormone actions are greatly amplified at the target cell. Activation of one receptor can activate many proteins.
Adenylate Cyclase-cAMP Polypeptide or glycoprotein hormone binds to receptor protein causing dissociation of subunit from G-protein complex. G-protein subunit binds to and activates adenylate cyclase. ATP cAMP + PPi cAMP attaches to the inhibitory subunit of protein kinase. Inhibitory subunit dissociates and activates protein kinase. Phosphorylates enzymes within the cell to produce hormone’s effects. Modulates activity of enzymes present in the cell. Alters metabolism of the cell. cAMP inactivated by phosphodiesterase.
Hormones That Bind to Nuclear Receptor Proteins Lipophilic steroid and thyroid hormones dissociate from carrier proteins. Diffuse through the target plasma membrane. Steroid receptors are located in cytoplasm and in the nucleus. Function within cell to activate genetic transcription. Messenger RNA directs synthesis of specific enzyme proteins that change metabolism. Each nuclear hormone receptor has 2 regions: A ligand (hormone)-binding domain. DNA-binding domain. Receptor must be activated by binding to hormone before binding to specific region of DNA called HRE (hormone responsive element). Located adjacent to gene that will be transcribed.
Other effects on hormone activity include: The concentration of a hormone in the blood is subject to control. It varies according to homeostatic need. The availability of a hormone to its receptor depends on the hormone’s rate of secretion, its rate of metabolic activation, the extend of its binding to plasma proteins if it is lipophilic, and its removal from the blood. Removal can be by metabolic inactivation or urinary excretion. Negative feedback maintains the plasma concentration of a hormone at a needed level. When a hormone’s concentration falls below a certain set point, the gland increases the secretion of the hormone. When the hormone’s level is above the set point, the secretion decreases. Many endocrine control systems involve neuroendocrine reflexes. Neural input to a gland regulates the gland’s secretion. The secretion rate of many hormones varies by a diurnal or circadian rhythm.
Endocrine disorders result from the hyposecretion or hypersecretion of a hormone. Factors producing hyposecretion include heredity, dietary deficiency, immunologic factors, and disease processes. Hyposecretion can be primary or secondary (due to the deficiency of the hormone’s tropic hormone). Replacement therapy of a hormone can often successfully treat the conditions from hyposecretion. Hypersecretion of a hormone can also be primary or secondary. Factors producing hypersecretion include tumors on the endocrine gland and immunologic factors. Endocrine dysfunction can also arise from the unresponsiveness of target cells to a hormone.
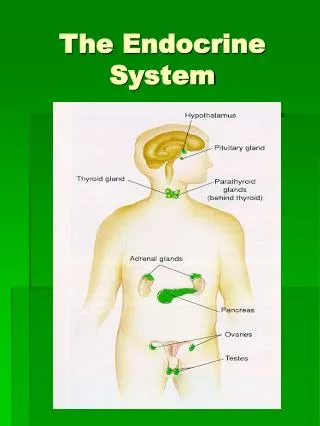
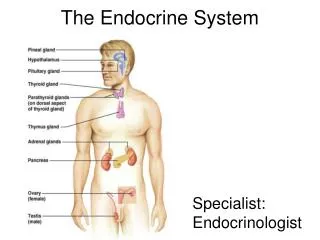
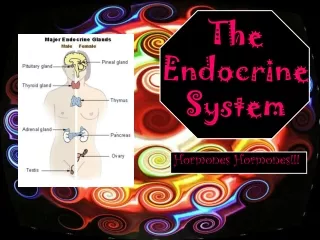
The pituitary gland is a small structure at the base of the brain. The posterior lobe is the neurohypophysis. It is composed of nervous tissue. The anterior lobe is the adenohypophysis. It is glandular tissue. The posterior lobe and the hypothalamus act as a unit to secrete vasopressin and oxytocin. The axons of the hypothalamus pass from the brain into capillaries in the posterior lobe. The posterior lobe does not produce vasopressin and oxytocin. They are produced by hypothalamic neurons. They are stored in neuron terminals in the posterior lobe. Vasopressin (ADH) signals the kidneys to retain water. It also signals the smooth muscle in the walls of arterioles. Its main role is regulating water balance.
The anterior pituitary secretes six hormones. Many are tropic. By a vascular network with the hypothalamus, each anterior pituitary hormone is secreted through signaling by a releasing hormone from this region of the brain. The thyroid-stimulating hormone (TSH) stimulates the secretion and growth of the thyroid gland. The adrenocorticotropic hormone (ACTH) stimulates the growth and secretion of hormones from the adrenal cortex. The follicle-stimulating hormone (FSH) stimulates growth and development of the ovarian follicles in females and sperm production in males. The luteinizing hormone (LH) stimulates ovulation and luteinization (female) and stimulates testosterone secretion in the male. Prolactin enhances breast development in females.
Hypothalamus Anterior pituitary Posterior pituitary TSH ACTH Prolactin Thyroid gland Adrenal cortex Mammary glands Thyroid hormone (T3 and T4) Cortisol Breast growth and milk secretion Metabolic actions; stress response Metabolic rate Fig. 17-6a, p. 539
Hypothalamus Anterior pituitary Posterior pituitary Growth hormone TSH ACTH or Many tissues Liver Metabolic actions Somatomedins Bone Soft tissues Growth Fig. 17-6b, p. 539
Hypothalamus Anterior pituitary Posterior pituitary Growth hormone TSH ACTH LH FSH Gonads (ovaries in females, testes in males) Sex hormone secretion (estrogen and progesterone in females, testosterone in males) Gamete production (ova in females, sperm in males) Fig. 17-6c, p. 539
Hypothalamic releasing and inhibiting hormones regulate anterior pituitary hormone secretion. TRH stimulates the release of TSH. CRH stimulates the release of ACTH. GnRH stimulates the release of FSH and LH. GHRH stimulates the release of the growth hormone. GHIH inhibits the release of the growth hormone and TSH. PRH stimulates the release of prolactin. PIH inhibits the release of prolactin. A hypothalamic hormone controls the output of an anterior pituitary hormone. The tropic hormone regulates the secretion of the target endocrine gland’s hormone.
The hypothalamic regulatory hormones reach the anterior pituitary by a vascular link. This is a capillary to capillary connection, the hypothalamic-hypophyseal portal system. Blood is this system carries hypothalamic signals to the anterior pituitary. Regulation of the secretion of the hypothalamic hormones depends on numerous inputs. Their complete regulation is not well understood. Target gland hormones inhibit hypothalamic and anterior pituitary hormone secretion via negative feedback. For example, a rise in cortisol from the adrenal cortex can feed back and reduce CRH secretion (hypothalamus) and the sensitivity of the ACTH secreting cells (anterior pituitary) to CRH. If cortisol falls in the blood, the direction of the other responses is reversed.
Neurosecretory neurons = Anterior pituitary hormone = Hypophysiotropic Hormones Hypothalamus Systemic arterial inflow Hypothalamic-hypophyseal portal system Anterior pituitary System venous outflow Posterior pituitary Fig. 17-8, p. 541
The endocrine system controls growth. Growth is signaled by the growth hormone. There are other factors that influence growth. Growth capacity is genetically determined. Adequate diet, freedom from chronic disease and stress, and normal levels of other growth-influencing hormones are other factors. The growth hormone does not play a role in fetal development. In children there is a postnatal growth spurt. The growth hormone may play a role in the later-occurring pubertal growth spurt. Androgens also contribute at this time.
The growth hormone has metabolic effects. It mobilizes fat stores (lipolytic) as a major energy source while conserving glucose for glucose-dependent tissues. This metabolic action is unrelated to growth. The growth hormone promotes growth by signaling an increase in the number of cells and size of cells in target organs. It stimulates the uptake of amino acids and protein synthesis in target cells. The growth hormone stimulates growth in the length and thickness of long bones. It stimulates the lengthening of bones at the epiphyseal plate. It stimulates osteoblast activity and the proliferation of epiphyseal cartilage. New bone tissue replaces cartilage in this region. It stimulates bone thickness by activating osteoblasts under the periosteum.
Hypothalamus Anterior pituitary Posterior pituitary Growth hormone TSH ACTH or Many tissues Liver Metabolic actions Somatomedins Bone Soft tissues Growth Fig. 17-6b, p. 539
The growth hormone exerts its effects indirectly by stimulating somatomedins. These substances are also called insulin-like growth factors. They are stimulated by the growth hormone and mediate most of the growth-promoting effects of the hormone. The main source of these factors is the liver. Their production depends on adequate nutrition. Their production is also related to age. The secretion of the growth hormone is regulated by GHRH and GHIH. Many factors influence the secretion of the growth hormone. It increases one hour after a deep sleep. Exercise can increase the secretion of the growth hormone. An abundance of amino acids and hypoglycemia increase its release.
Blood amino acids, Blood fatty acids Exercise, stress, blood glucose Hypothalamus Diurnal rhythm Growth hormone– inhibiting hormone (GHIH) Growth hormone– releasing hormone (GHRH) Anterior pituitary Growth hormone Liver Metabolic actions unrelated to growth: fat breakdown ( blood fatty acids) glucose uptake by muscles ( blood glucose) Somatomedins Growth-promoting actions: cell division protein synthesis ( blood amino acids) bone growth Fig. 17-10, p. 545
A deficiency or excess of the growth hormone changes growth patterns. A hyposecretion produces dwarfism in a child. In Laron dwarfism, tissues fail to respond to the growth hormone. In adults a growth hormone deficiency reduces muscle mass and strength. A hypersecretion of the growth hormone produces gigantism in the child. If hypersecretion occurs after the epiphyseal plates have closed, acromegaly develops. Only certain bones are affected. Other hormones in addition to the growth hormone are essential for normal growth. The thyroid hormone is essential for growth. Insulin is a growth promoter. Androgens play a role in a pubertal growth spurt.
The pineal gland is a small structure in the brain. It secretes the hormone melatonin. It helps keep the body’s circadian rhythms in synchrony with the light-dark cycle. The suprachiasmatic nucleus (SCN) has a major role in establishing many of the body’s daily rhythms. It secretes clock proteins. Cyclic changes in their concentration changes the neural output from the SCN. The SCN works in conjunction with the pineal gland and pineal gland to regulate circadian rhythms. Daily changes in light intensity is the major environmental factor used to adjust the SCN master clock. Melatonin has other functions not related to circadian timekeeping. It accomplishes natural sleep without hypnosis (side effects). It inhibits hormones that stimulate reproductive activity. It is also an effective antioxidant.
The thyroid gland consists of two lobes of endocrine tissue. It lies over the trachea, below the larynx. Its follicular cells store colloid. Thyroglobulin (TGB) is the main constituent of this colloid. The follicular cells produce two hormones, T4 and T3. These two hormones are collectively the thyroid hormone. It regulates overall basal metabolic rate. C cells between the follicular cells secrete calcitonin. It plays a role in calcium metabolism. Tyrosine and iodine are the ingredients for the thyroid hormone. The thyroid hormone synthesis occurs on the thyroglobulin molecules in the colloid.
Colloid Blood Thyroid follicular cell *Endoplasmic reticulum/Golgi complex Lysosome DIT = Di-iodotyrosine T3 = Tri-iodothyronine T4 = Tetraiodothyronine (thyroxine) TGB = Thyroglobulin I = Iodine MIT = Monoiodotyrosine Fig. 17-14, p. 549
Thyroid hormone synthesis and storage occurs through a series of steps. Tyrosine is incorporated into TGB. This is transported into the colloid by exocytosis. Iodine is transported into the colloid by follicular cells. The attachment of one iodine to tyrosine produces MIT. The attachment of two iodines to tyrosine produces DIT. The coupling of two DITs produces T4. The coupling of one MIT to two DITS produces T3. Thyroid follicular cells engulf a part of TGB-containing colloid by phagocytosis. Lysosomes attack the engulfed vesicle and split the iodinated products from TGB. T4 and T3 reach the blood by diffusion. The deiodination of T4 and T3 free the iodine for recycling. The thyroid hormone is highly lipophilic. It binds to several plasma proteins.
Most of the T4 is converted to T3 outside the thyroid. The thyroid hormone has many metabolic effects. T4 loses one of it iodines in the liver or kidney. The hormone increases the body’s overall basal metabolic effect. It regulates the body’s use of oxygen and is calorigenic (heat-producing). Large amounts of the secreted hormone convert glycogen into glucose and stimulates protein degradation. This hormone also has sympathomimetic effects, increasing target cells’ responsiveness to epinephrine and norepinephrine. It increases heart rate and the force of heart contraction. It also stimulates growth hormone secretion and promotes the effect of this hormone on increased protein synthesis.
Cold in infants Stress Hypothalamus Thyrotropin- releasing hormone (TRH) Anterior pituitary Thyroid-stimulating hormone (TSH) Thyroid gland Thyroid hormone (T3 and T4) Metabolic rate and heat production; enhancement of growth and CNS development; enhancement of sympathetic activity Fig. 17-15, p. 551
The secretion of the thyroid hormone is regulated by the hypothalamus-pituitary-thyroid axis. TSH from the anterior pituitary stimulates the release of the thyroid hormone. TSH also maintains the structural integrity of the thyroid gland. TRH from the hypothalamus turns on TSH secretion. An increase in the thyroid hormone feeds back to decrease TSH secretion (negative feedback). TRH secretion is increased only by exposure to the cold in newborn infants.
Imbalances in the thyroid hormone cause changes in development. Hypothyroidism produces myxedema in the adult. From birth a deficiency of the hormone produces cretinism. Causes of these conditions include deficiency of TRH, TSH, or the thyroid hormone. A deficiency of iodine in the diet can also be a cause. Symptoms include a lowered basal metabolic rate, excessive weight gain, bradycardia, cold intolerance and the quick onset of fatigue. Grave’s disease is the most common cause of hyperthyroidism. It is an autoimmune disease. Symptoms include an elevated metabolic rate, high heart rate, heat intolerance and exophthalmos. A goiter develops when the thyroid gland is overstimulated. Hypothyroidism leads to high levels of TSH due to only a small amount of negative feedback. TSH acts on the follicular cells to increase their size and number. A goiter can develop in Grave’s disease due to an increase in thyroid-stimulating immunoglobulin (TSI).
Anterior pituitary Thyroid-stimulating immunoglobulin (TSI) No TSH (No stimulation) Thyroid gland Thyroid hormone Fig. 17-16, p. 551
There are two adrenal glands. Each is embedded in a capsule of fat on top of each kidney. The outer adrenal cortex of each gland secretes several steroid hormones. The inner adrenal medulla of each gland secretes epinephrine and norepinephrine. The adrenal cortex consists of three different zones. Each secretes a different family of hormones. One of these zones secretes the mineralocorticoids. The mineralocorticoids (e.g., aldosterone) signal the kidneys (distal tubule and collecting duct) to retain sodium (plus water) and eliminate potassium. Aldosterone secretion is increased by activation of the renin-angiotensin-aldosterone system.
Diurnal rhythm Stress Hypothalamus Corticotropin-releasing hormone (CRH) Anterior pituitary Adrenocorticotropic hormone (ACTH) Adrenal cortex Cortisol Blood glucose (by stimulating gluconeogenesis and inhibiting glucose uptake) Metabolic fuels and building blocks available to help resist stress Blood amino acids (by stimulating protein degradation) Blood fatty acids (by stimulating lipolysis) Fig. 17-20, p. 555
One of the zones in the adrenal cortex secretes the glucocorticoids. The glucocorticoids (mainly cortisol) stimulate gluconeogenesis. This is the conversion of amino acids into carbohydrates, occurring mainly in the liver. Cortisol also facilitates the inhibition of glucose uptake, stimulates protein degradation, and promotes lipolysis. Cortisol plays a major role in the adaptation to stress. Noxious stimuli that can produce this include physical, chemical, physiologic, psychological, emotional, and social sources. An increased concentration of glucose in the blood is major response to all of these. Pharmacologic levels of cortisol can have anti-inflammatory and immunosuppressive effects. This can be used to treat rheumatoid arthritis or allergies. Long-term use of this treatment can produce unwanted side effects.
The secretion of cortisol is regulated by the hypothalamus-pituitary-adrenal cortex axis. ACTH from the anterior pituitary stimulates the secretion of cortisol from the adrenal cortex. ACTH secretion is triggered by CRH from the hypothalamus. Negative feedback from cortisol in the blood to the hypothalamus and the anterior pituitary regulate the level of cortisol in the blood. Increased output of CRH and ACTH increases in response to stress. Cortisol secretion also varies by a diurnal rhythm.
A third zone in the adrenal cortex secretes androgens or estrogens. Both are produced in either sex. Usually they are not abundant enough to be powerful in either sex. The androgens have masculinizing effects. The androgen DHEA can have an effect in females who otherwise lack androgens. ACTH controls adrenal androgen secretion.