Download

1 / 51
510 likes | 594 Views
Learn about manipulating DNA in the lab, nucleic acid synthesis, solid-phase synthesis, cloning, plasmids, and more in recombinant DNA technology. Discover how to build useful plasmids and vectors for genetic manipulation.

E N D
Recombinant DNA Andy HowardIntroductory Biochemistry29 October 2014 Recombinant DNA I
DNA can be manipulated • We’ll show how DNA is replicated in the laboratory and we’ll explore some specific aspects of recombinant DNA technology and its applications Recombinant DNA I
What we’ll discuss • DNA synthesis • Solid-phase synthesis • Phosphoramidates • Cloning • Probes • Hybridization, blots • High-throughput techniques • Prokaryotic Expression • Eukaryotic Expression Recombinant DNA I
Synthesizing nucleic acids • Laboratory synthesis of nucleic acids requires complex strategies • Functional groups on the monomeric units are reactive and must be blocked • Correct phosphodiester linkages must be made • Recovery at each step must high! Recombinant DNA I
Solid Phase Oligonucleotide Synthesis • Dimethoxytrityl group blocks the 5'-OH of the first nucleoside while it is linked to a solid support by the 3'-OH • Step 1:Detritylation by trichloroacetic acid exposes the 5'-OH • Step 2: In coupling reaction, second base is added as a nucleoside phosphoramidate Recombinant DNA I
Synthesis I • Figure 11.29Solid phase oligonucleotide synthesis. The four-step cycle starts with the first base in nucleoside form (N-1) attached by its 3'-OH group to an insoluble, inert resin or matrix, typically either controlled pore glass (CPG) or silica beads. Its 5'-OH is blocked with a dimethoxytrityl (DMTr) group (a). Recombinant DNA I
Blocking groups If the base has reactive -NH2 functions, as in A, G, or C, then N-benzoyl or N-isobutyryl derivatives are used to prevent their reaction (b). In step 1, the DMTr protecting group is removed by trichloroacetic acid treatment. Step 2 is the coupling step: the second base (N-2) is added in the form of a nucleoside phosphoramidite derivative whose 5'-OH bears a DMTr blocking group so it cannot polymerize with itself (c). Recombinant DNA I
Solid Phase Synthesis • Step 3: capping with acetic anhydride blocks unreacted 5’-OHs of N-1 from further reaction • Step 4: Phosphite linkage between N-1 and N-2 is reactive and is oxidized by aqueous iodine to form the desired, and more stable, phosphate group Recombinant DNA I
Activation of the phosphoramidate Recombinant DNA I
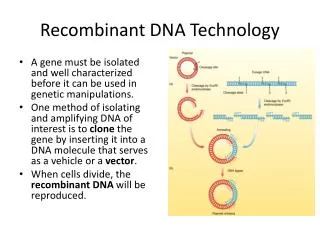
Cloning • Cloning is the process whereby DNA is copied in a controlled way to produce desired genetic results Recombinant DNA I
Plasmids • Small (typically < 10 kbp), usually circular segments of DNA that get replicated along with the organism’s chromosome(s) • Bacterial plasmids have a defined origin of replication and segments defining specific genes • Some are natural;others are man-made Recombinant DNA I
How they’re used • Typical man-made plasmid includes a gene that codes for an enzyme that renders the bacterium resistant to a specific antibiotic, along with whatever other genetic materials the experimenter or clinician wishes to incorporate • Thus the cells that have replicated the plasmid will be antibiotic-resistant; surviving colonies will be guaranteed (?) to contain the desired plasmid in all its glory Recombinant DNA I
A typical plasmid Recombinant DNA I
Building useful plasmids • Take starting plasmid and cleave it with a restriction enzyme at a specific site • Add foreign DNA that has been tailored to fit into that plasmid Recombinant DNA I
Inserts • Typically a place within the plasmid will be set up so that small stretches (< 10 kbp) of desired DNA can be ligated in • With sticky ends: high specificity, but you do get self-annealing of the plasmid and of the insert, so those have to be eliminated • With blunt ends: require more artisanry:T4 phage ligase can rejoin ends without stickiness; but it’s chaotic Recombinant DNA I
Directional cloning • Guarantees that the desired DNA goes in in exactly one orientation Recombinant DNA I
Use of bacteriophage lambda • Can handle somewhat larger inserts (10-16 kbp) • Middle third of its 48.5-kbp chromosome isn’t needed for infection Recombinant DNA I
Cosmids • 14-bp sequence cos (cohesive end site):5’-TACGGGGCGGCGACCTCGCG-3’3’-ATGCCCCGCCGCTGGAGCGC-5’ • … one of these at each end • Must be 36 kbp < separation < 51 kbp apart • In practice we can use these for inserts up to 40 kbp in size Recombinant DNA I
Cosmids in action(fig. 12.9) Recombinant DNA I
Shuttle vectors • These are plasmids that can operate in two different organisms • Usually one prokaryote and one eukaryote (e.g. Escherichia coli and Saccharomyces cerevisiae) • Separate origins for each host • This allows us to clone the vector in a bacterial host and then express it in a eukaryotic setting Recombinant DNA I
Typical shuttle vector Recombinant DNA I
Artificial chromosomes • Huge chunks (2 megaBp!) can be propagated in yeast with artificial chromosomes (YACs) • These can be manipulated in the yeast setting or transferred to transgenic mice in a living animal • YACs need origin, a centromere, and telomeres Recombinant DNA I
Use of YACs in mice • Diagram courtesy Expert Reviews in Molecular Medicine, 2003 Recombinant DNA I
DNA libraries • Set of cloned fragments that make up an organism’s DNA • We can isolate genes from these • Most common approach to creating these is shotgun cloning, in which we digest the total DNA and then clone fragments into vectors • Goal is that ≥ 1 clone will contain at least part of the gene of interest (might have been clipped by the restriction enzyme!) Recombinant DNA I
Probabilities • Probability P that some number of clones, N, contains a particular fragment representing a fraction f of the genome:P = 1 - (1 - f)N • Therefore 1-P = (1-f)N • Thus ln(1-P) = ln{(1-f)N} = Nln(1-f) • Therefore N = ln(1-P) / ln(1-f) Recombinant DNA I
What that means • The value f is pretty small, so the denominator is only slightly negative; whereas we want the numerator to be very negative, since that corresponds to a high value of P. • 10 kbp fragments in E.coli meansf = 10/4640 = 0.0022,so for P = 0.99, we need N=1.4*106 • We’d do better with larger f values! Recombinant DNA I
Finding relevant fragments by colony hybridization • Plate out a library of fragments and grow colonies or plaques • Soak those onto a flexible absorbent disc • Disc is treated with high-pH to dissociate bound DNA duplexes; placed in a sealed bag with a radiolabeled probe • If they hybridize, radioactivity will stick to disc • The hits can be recovered from the master plate Recombinant DNA I
Colony hybridization illustrated Recombinant DNA I
Making the probes • Sometimes we have at least part of the gene sequence and can fish for it • Other times we know the amino acid sequence and can work backward, but with degeneracy(64 codons, 20 aa’s) • Typically use at least 17mers to guarantee that they don’t get random association • Probes derived from a different species are heterologous • With big eukaryotic genes we may have to look for pieces of the gene, not the whole thing Recombinant DNA I
cDNA libraries • Sometimes the easiest thing to obtain are mRNA templates associated with a particular function • Reverse transcriptase can make a complementary (cDNA) molecule from such an mRNA template • A library of cDNAs can be assembled from a collection of mRNA templates Recombinant DNA I
Why is that useful? • The mRNAs will be unique to the cell type from which they’re derived • Often they’re also unique to the functional role that tissue is playing at the time • Therefore finding that collection of cDNAs tells us about cellular activity Recombinant DNA I
Expressed sequence tags • An EST is a short (~200 base) sequence derived from a cDNA • Represents part of a gene that is being expressed • Labeled ESTs can be mounted on a gene chip and used to identify cells that are expressing a particular class of mRNAs Recombinant DNA I
Southern blots I: fractionation • Tool for identifying a particular DNA fragment out of a vast population thereof • Exploits sequence specificity for identification • Developed by E.M.Southern in 1975 • Begins with electrophoretic fractionation of fragments (mobility 1/mass) • Polyacrylamide gels ok 25-2000 bp; agarose better for larger fragments Recombinant DNA I
Southern blots 2: blotting • Gel soaked in base to denature duplexes • pH readjusted to neutral • Sheet of absorbent material placed atop the gel • Salt solution is drawn across the gel, perpendicular to the electrophoretic direction, in various ways to carry the DNA onto the sheet • Sheet is dried in an oven to tightly attach the DNA to it • Incubate sheet with protein or detergent to saturate remaining DNA binding sites on sheet so we don’t get nonspecific binding Recombinant DNA I
Southern blots 3: hybridization • Labeled probe and sheet placed in sealed bag • If probe attaches, label will appear at that point on the sheet via annealing or hybridization • Label detected by autoradiography Recombinant DNA I
Southern blots illustrated Recombinant DNA I
Variations on this idea • RNA can be used as the probe: that’s called a Northern blot • Proteins can be substituted by using an antibody as a probe and a collection of protein fragments as the analytes; that’s called a Western blot • Various groups have tried to coin the term Eastern blot, but no agreement has been reached as to what that would be (aptamer? Lectin?) Recombinant DNA I
High-throughput techniques • Eagerness to provide rapid, easy-to-use applications of these approaches has led to considerable research on ways to make these techniques work fast and automatically • This high-throughput approach enables many experiments per unit time or per dollar Recombinant DNA I
DNA microarrays • Thousands of oligonucleotides immobilized on a substrate • Synthesis by solid-phase phosphoramidite chemistry • Typically 25-base oligos • Can be used in cDNA projects to look at expression patterns Recombinant DNA I
An example Recombinant DNA I
Using expression vectors • We often want to do something with cloned inserts in expression vectors, viz. make RNA or even protein from it • RNA: stick an efficient promoter next to the cloning site; vector DNA transcribed in vitro using SP6 RNA polymerase • This can be used as a way of making radiolabeled RNA Recombinant DNA I
Protein expression • Making (eukaryotic) proteins in bacteria via cDNA means we don’t have to worry about introns • Expression vector must have signals for transcription and translation • Sequence must start with AUG and include a ribosome binding site • Strong promoters can coax the bacterium into expressing 30% of E.coli’s protein output to be the one protein we want! Recombinant DNA I
Example: ptac • This is a fusion of lac promoter (lactose metabolism) with trp promoter (tryptophan biosynthesis) • Promoter doesn’t get turned on until an inducer (isopropyl--thiogalactoside, IPTG) is introduced Recombinant DNA I
iClicker quiz, question 1 1. Probe systems employing RNA are called • (a) Southern blots • (b) Northern blots • (c) Western blots • (d) Eastern blots • (e) None of the above Recombinant DNA I
iClicker quiz, question 2 2. The inducer used with the ptac promoter system is • (a) glucose • (b) glucose-6-phosphate • (c) IPTG • (d) ionizing radiation • (e) none of the above. Recombinant DNA I
Eukaryotic expression • Sometimes we need the glycosylations and other PTMs that eukaryotic expression enables • This is considerably more complex • Common approach is to use vectors derived from viruses and having the vector infect cells derived from the virus’s host • Example: baculovirus, infecting lepidopteran cells; gene cloned just beyond promoter for polyhedrin, which makes the viral capsid protein Recombinant DNA I
Screening libraries with antibodies • Often we have antibodies that react with a protein of interest • If we set up a DNA library and introduce it into host bacteria as in colony hybridization, we can put nylon membranes on the plates to get replicas of the colonies • Replicas are incubated to make protein • Cells are treated to release the protein so it binds to the nylon membrane • If the antibody sticks to the nylon, we have a hit! Recombinant DNA I
Fusion proteins • Sometimes it helps to co-express our protein of interest with something that helps expression, secretion, or behavior • We thereby make chimeric proteins, carrying both functionalities • We have to be careful to keep the genes in phase with one another! • Often the linker includes a sequence that is readily cleaved by a commercial protease Recombinant DNA I
Fusion systems (table 12.2+) Recombinant DNA I
Improving purification via expression • If we attach (usually at the N-terminal end) a his-tag (several his, several cys) to our protein, it becomes easier to purify: • The his tag forms a loop that will bind strongly to a divalent cation like Ni2+ • Thus we can pour our expressed protein through a Ni2+ affinity column and it will stick, while other proteins pass through • We elute it off by pouring through imidazole, which completes for the Ni2+ and lets our protein fall off Recombinant DNA I