Download

1 / 22
260 likes | 740 Views
Chapt. 9 Regulation of Enzymes. Regulation of Enzymes Student Learning Outcomes : Explain that enzyme activities must be regulated for proper body function Explain three general mechanisms: Reversible binding in active site: substrate, inhibitors

E N D
Chapt. 9 Regulation of Enzymes • Regulation of Enzymes • Student Learning Outcomes: • Explain that enzyme activities must be regulated for proper body function • Explain three general mechanisms: • Reversible binding in active site: • substrate, inhibitors • Changing conformation of active site of enzyme: • Allosteric effectors, covalent modification, • Protein-protein interactions, zymogen cleavage • (Changing concentration of enzyme) • Synthesis, degradation
Regulation of metabolic pathways • Metabolic pathway analogous to cars on highway: • Flux of substrates affected by rate-limiting enzyme (barrier) • Removal of barrier increases flow • Activating rate-limiting enzyme Fig. 9.1
Regulation of glucose metabolism pathway • Ex. Regulation of glucose metabolism pathway: • Hexokinase & glucokinases convert glucose -> G-6-P in cells • Glycolysis for energy • Feedback regulation by ATP • Store G-6-P as glycogen • Feedforward by insulin
II. Regulation by substrate, product concentration • Michaelis-Menten equation describes kinetics: • More substrate gives more reaction, to maximal • Vi (initial velocity) relates to concentration of substrate [S] to Vmax (maximal velocity) and Km ([S] for 1/2 Vmax • Applies to simple reactions: • E + S ES E + P; k1 = forward, k2 back; k3 for E+P • Vi = Vmax[S]/ Km + [S] Km = k2 + k3/k1; Vmax = k3[Et]
II. Regulation by substrate, product concentration: • Ex.Graph of Michaelis-Menten equation has limit of Vmax at infinite substrate. • Km = [S] where Vmax/2 • Ex. Glucokinase Km 5 mM: • If blood glucose 4 mM -> • Vi = 0.44 Vmax • (Vm x 4mM/ (5mM + 4 mM) • Blood glucose 20 mM -> • Vi = 0.8 Vmax • (Vm x 20mM/ 5 + 20 mM Fig. 9.2
Different isozymes have different Km for glucose • Different hexokinases differ in Km for glucose: • glucose + ATP -> G-6-P + ADP • Hexokinase I • (rbc) only glycolysis • Glucokinase • (liver, pancreas) storage • Fasting blood sugar • about 5 mM (90 mg/dL) so • rbc can function even if low • blood sugar of glucose • S0.5 = half-max for S-shape curve Fig. 9.3
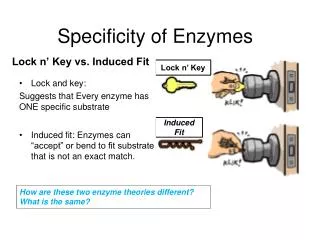
Reversible inhibitors decrease reaction velocity • Regulation through active site: reversible inhibitors • Competitive inhibitors compete with substrate Overcome by excess substrate (increase apparent Km) • Noncompetitive do not compete with substrate Not overcome by substrate (lowers [E] and Vmax) Products can also inhibit enzyme activity Fig. 9.4
III. Regulation through conformational changes • Regulation through conformational changes of enzyme can affect catalytic site: • Allostery • – ex. Glycogen phosphorylase • Phosphorylation • – ex. Glycogen phosphorylase kinase • Protein-protein interactions • - ex. Protein kinase A • Proteolytic cleavage • - ex. chymotrypsinogen
A. Allosteric Activators and inhibitors • Allosteric enzymes: • Often multimeric, • Exhibit positive cooperativity in substrate binding (ex. Hemoglobin and O2) • T (taut state) inactive without substrate • R (relaxed) state active with substrate Fig. 9.5
Allosteric activators and inhibitors • Allosteric enzymes often cooperative S binding • Allosteric activators and inhibitors: • Bind at allosteric site, • not catalytic site • Conformational change • Activators often bind • R (relaxed) state • decrease S0.5 • Inhibitors often bind • T (taut state) • increase S0.5 Fig. 9.6
B. Conformational change by covalent modification • Phosphorylation can activate or inhibit enzymes: • Protein kinases add phosphate • Protein phosphatases remove • PO42- adds bulky group, • negative charge, interacts • with other amino acids Fig. 9.7
Muscle glycogen phosphorylase regulation • Muscle glycogen phosphorylase is regulated by both phosphorylation and/or allostery: • Rate-limiting step glycogen -> glucose-1-PO4 • ATP use increases AMP - allostery • phosphorylation increases activity • Signal from PKA Fig. 9.8
Ex. Protein kinase A • Protein kinase A: Regulatory, catalytic subunits: • Ser/thr protein kinase, phosphorylates many enzymes • Including glycogen phosphorylase kinase • Adrenline increase cAMP, dissociates R subunits, • Starts PO4 cascade Fig. 9.9 cAMP activates PKA
Other covalent modifications affect proteins • Covalent modifications affect protein activity, location in cell: • acetyl- (on histones) • ADP-ribosylation • (as by cholera toxin on Ga subunit) • Lipid addition (as on Ras protein) Fig. 6.13 modified amino acids
Conformational changes from Protein-Protein interactions • Ca-Calmodulinfamily of modulator proteins • activated when [Ca2+ ] increases. • Ca2+/calmodulin binds to targets • e.g. protein kinases, allosteric result CaMkinasefamily activated by Ca2+/calmodulin; phosphorylate metabolic enzymes, ion channels, transcription factors, regulate synthesis, release of neurotransmitters. Fig. 9.10
Small monomeric G proteins • Small (monomeric) G proteins • affect conformation of other proteins: • GTP bound form binds and activates or inhibits • GDP bound form inactive • Other intermediates regulate the G proteins (GEF, GAP, etc) • Ras family (Ras, Rho, Rab, Ran, Arf) • diverse roles in cells Fig. 9.11
Proteolytic cleavage is irreversible • Proteolytic cleavage is irreversible conformational change: • Some during synthesis and processing • Others after secretion: • Proenzymes inactive: • Ex. Precursor protease is zymogen: • (chymotrypsinogen is cleaved by trypsin in intestine) • Ex. Blood clotting factors fibrinogen, prothrombin
Regulation of pathways • Regulation of metabolic pathways is complex: • Sequential steps, different enzymes, rate-limiting one • Match regulation to function of path Fig. 9.12
Lineweaver-Burk plot • Lineweaver-Burk transformation converts Michaelis-Menten to straight line (y = mx + b) • double reciprocal plot • Ease of determining • Km and Vmax Fig. 9.13
Lineaver-Burk plots permit comparisons • Lineweaver-Burk plots permit analysis of enzyme kinetics, characterization of inhibitors Fig. 9.14
Key concepts • Key concepts: • Enzyme activity is regulated to reflect physiological state • Rate of enzyme reaction depends on concentration of substrate, enzyme • Allosteric activators or inhibitors bind sites other than the active site: conformational • Mechanisms of regulation of enzyme activity include: feedback inhibition, covalent modifications, interactions of modulator proteins (rate synthesis, degradation)
Review questions • 3. Methanol (CH3OH) is converted by alcohol dehydrogenases (ADH) to formaldehyde (CH2O), a highly toxic compound . Patients ingested toxic levels of methanol can be treated with ethanol (CH3CH2OH) to inhibit methanol oxidation by ADH. Which is the best rationale for this treatment? • Ethanol is structural analog of methanol – noncompetitive inhibitor • Ethanol is structural analog of methanol – will compete with methanol for binding enzyme • Ethanol will alter the Vmax of ADH for oxidation of methanol. • Ethanol is effective inhibitor of methanol oxidation regardless of the concentration of methanol • Ethanol will inhibit enzyme by binding the formadehyde-binding site on the enzyme, even though it cannot bind the substrate binding site for methanol.