
Citric Acid Cycle: Key Reactions, Energy Production, and Vitamin Roles
300 likes | 465 Views
Learn about the citric acid cycle, its key reactions, energy production mechanism, and the essential roles of B vitamins in metabolism. Explore how this cycle plays a crucial role in gluconeogenesis, transamination, and deamination processes.

Citric Acid Cycle: Key Reactions, Energy Production, and Vitamin Roles
E N D
Presentation Transcript
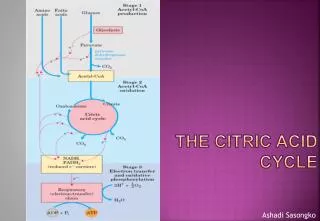
Citric Acid Cycle • The citric acid cycle is the final common pathway for the oxidation of carbohydrate, lipid, and protein because glucose, fatty acids, and most amino acids are metabolized to acetyl-CoA or intermediates of the cycle. It also has a central role in gluconeogenesis, lipogenesis, and interconversion of amino acids. Many of these processes occur in most tissues, but the liver is the only tissue in which all occur to a significant extent.
Citric Acid Cycle is also called as the tricarboxylic acid cycle or krebes cycle. It is a process of the oxidation of acetyl CoA, to CO2 and H2O • Reactions of Citric Acid Cycle • Citrate synthase: • In the first step, acetyl CoA (formed from pyrovate under aerobic condition)combines with oxaloacetate and formed Citric Acid (a tricarboxylic acid).
2. Aconitase: This enzyme catalyses the isomerization reaction by removing and then adding back the water ( H and OH ) to cis-aconitate in at different positions. Both the steps are carried out by Aconitase .Conversion of citrate to isocitrate is inhibited by fluoroacetate.
3. Isocitrate dehydrogenase: In the presence of Isocitratedehydrogenase, isocitrate is converted to oxalosuccinate which is later decarboxylated to form α-ketoglutarate
4. α-Ketoglutarate dehydrogenase: This is a complex of different enzymatic activities similar to the pyruvate dehydrogenase complex. It has the same mechanism of reaction with E1, E2 and E3 enzyme unites, and requires five coenzyme,i.e. TPP, NAD+ ,FAD, coenzyme A and Lipoic acid.
5. Succinate thiokinase: Sccinyl CoA is changed to succinate by succinate thiokinase. during this reaction, amolecule of GTP is formed. This in turn is equivalent to ATP and is known as substrate level phosphorylation since a high energy molecule is formed at the substrate level only. Succinate thiokinase
6. Succinate Dehydrogenase: Oxidation of succinate to fumarate.This is the only citric acid cycle enzyme that is tightly bound to the inner mitochondrial membrane. It is an FAD dependent enzyme. Due to structural similarity between malonate and succinate, malonate inhibits succinate dehydrogenase, competitively.
7. Fumarase: Hydration of Fumarate to malate: It is a highly stereospecific enzyme.
8. L-Malate dehydrogenase: Oxidation of malate to oxaloacetate: It is an NAD+ dependent enzyme.
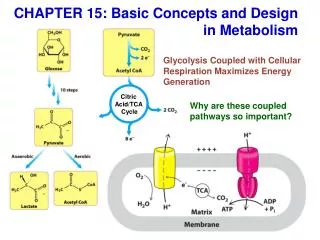
Energy production during krebs cycle As a result of the oxidation of one molecule of acetyl CoA in the krebs cycle, three molecules of NAD+ and one molecule of FAD are reduced. Reducing equivalents(from NADH+H+) enters the respiratory chain and results in the production of three molecules of ATP. Similarly , FADH2 yields 2ATP. Besides, there is also a substrate level production of GTP. Total ATP yield, per molecule of acetyl CoA , is thus 12 ATP(Table 1.1)
Table(1.1) Energy production from Citric Acid Cycle Isocitate α-ketoglutarate NADH+H+ (3ATP) α-ketoglutarate Succinyl CoA NADH+H+ (3ATP) Succinyl CoA Succinate - (1ATP) Succinate Fumarate FADH2 (2ATP) Malate Oxaloacetate NADH+H+ (3ATP) total (12ATP) In addition, as mentioned above, conversion of pyruvate to acetyl CoA also generates NADH+H+ and gives 3ATP
Thus, the total number of ATP produced from the oxidation of pyruvate is 15 Since 2 molecules of pyruvate are formed from one molecule of glucose, therefore, in addition to energy yield during aerobic glycolysis (8ATP) a molecule of glucose produces 30 ATP via Krebs cycle. Thus, a total of 38 ATP are obtained when a molecule of glucose is completely oxidized to CO2 and H2O under aerobic condition, i.e. via glycolysis and Krebs cycle.
VITAMINS PLAY KEY ROLES IN THE CITRIC ACID CYCLE Four of the B vitamins are essential in the citric acid cycle and hence energy-yielding metabolism: (1)B2 (riboflavin), in the form of flavin adenin dinucleotide (FAD), a cofactor for succinate dehydrogenase; (2) niacin,in the form of nicotinamide adenine dinucleotide (NAD), the electron acceptor for isocitrate dehydrogenase, α-ketoglutarate dehydrogenase, and malate dehydrogenase; (3) thiamin(vitamin B1 ), as thiamin pyrophosphate(TPP), the coenzyme for decarboxylation in the α-ketoglutarate dehydrogenase reaction; and (4)pantothenic acid, as part of coenzyme A, the cofactor attached to "active" carboxylic acid residues such as acetyl-CoA and succinyl-CoA.
THE CITRIC ACID CYCLE PLAYS A PIVOTAL ROLE IN METABOLISM The citric acid cycle is not only a pathway for oxidation of two-carbon units, but is also a major pathway for interconversion of metabolites arising from transamination and deamination of amino acids and providing the substrates for amino acid synthesis by transamination as well as for gluconeogenesis and fatty acid synthesis. Because it functions in both oxidative and synthetic processes, it is amphibolic.
The Citric Acid Cycle Takes Part in Gluconeogenesis, Transamination, &Deamination All the intermediates of the cycle are potentially glucogenic, since they can give rise to oxaloacetate, and hence net production of glucose (in the liver and kidney, the organs that carry out gluconeogenesis. The key enzyme that catalyzes net transfer out of the cycle into gluconeogenesis is phosphoenolpyruvate carboxykinase, which catalyzes the decarboxylation of oxaloacetate to phosphoenolpyruvate, with GTP acting as the phosphate donor. Net transfer into the cycle occurs as a result of several reactions. Among the most important of such anaplerotic reactions is the formation of oxaloacetate by the carboxylation of pyruvate, catalyzed by pyruvatecarboxylase.
This reaction is important in maintaining an adequate concentration of oxaloacetate for the condensation reaction with acetyl-CoA. If acetyl-CoA accumulates, it acts as both an allosteric activator of pyruvate carboxylase and an inhibitor of pyruvate dehydrogenase, thereby ensuring a supply of oxaloacetate. Lactate, an important substrate for gluconeogenesis, enters the cycle via oxidation to pyruvate and then carboxylation to oxaloacetate.
Aminotransferase (transaminase) reactions form pyruvate from alanine, oxaloacetate from aspartate, and α-ketoglutarate from glutamate. Because these reactions are reversible, the cycle also serves as a source of carbon skeletons for the synthesis of these amino acids.
Other amino acids contribute to gluconeogenesis because their carbon skeletons give rise to citric acid cycle intermediates. Alanine, cysteine, glycine, hydroxyproline, serine, threonine, and tryptophan yield pyruvate; arginine, histidine, glutamine, and proline yield α-ketoglutarate; isoleucine, methionine, and valine yield succinyl-CoA; tyrosine and phenylalanine yield fumarate
The Citric Acid Cycle Takes Part in Fatty Acid Synthesis Acetyl-CoA, formed from pyruvate by the action of pyruvate dehydrogenase, is the major substrate for long-chain fatty acid synthesis in nonruminants. (In ruminants, acetyl-CoA is derived directly from acetate.) Pyruvate dehydrogenase is a mitochondrial enzyme, and fatty acid synthesis is a cytosolic pathway; the mitochondrial membrane is impermeable to acetyl-CoA. Acetyl-CoA is made available in the cytosol from citrate synthesized in the mitochondrion, transported into the cytosol, and cleaved in a reaction catalyzed by ATP-citrate lyase.
Citrate is only available for transport out of the mitochondrion when aconitase is saturated with its substrate, and citrate cannot be channeled directly from citrate synthase onto aconitase. This ensures that citrate is used for fatty acid synthesis only when there is an adequate amount to ensure continued activity of the cycle.
Regulation of the Citric Acid Cycle Citrate synthase, isocitrate dehydrogenase and α-ketoglutarate dehydrogenase are rate limiting enzymes of the cycle. NADH, ATP and succinyl CoA function as inhibitors while ADP and Ca+2 are activators of the cycle.














![Tricarboxylic acid cycle (TCA Cycle) [Kreb’s cycle] [Citric acid cycle]](https://cdn3.slideserve.com/6696193/tricarboxylic-acid-cycle-tca-cycle-kreb-s-cycle-citric-acid-cycle-dt.jpg)


