Download

1 / 14
190 likes | 814 Views
Proteasome & other proteases. 19-1. Proteasome - core complex and regulatory cap Other proteases - HslUV, ClpAP, ClpXP, Lon, FtsH. The proteasome. eukaryotic proteasome seven types of alpha subunits seven types of beta subunits regulatory cap (at least 17 subunits).

E N D
Proteasome & other proteases 19-1 Proteasome - core complex and regulatory cap Other proteases - HslUV, ClpAP, ClpXP, Lon, FtsH
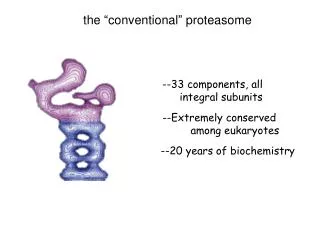
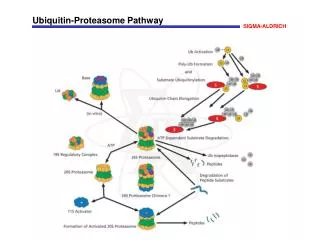
The proteasome • eukaryotic proteasome • seven types of alpha subunits • seven types of beta subunits • regulatory cap (at least 17 subunits) core particle (20S) 26S regulatory particle (19S) 19-2 • the proteasome is a cylindrical proteasome consisting of four stacked, seven-membered rings • the two outer rings are alpha subunits (inactive) • the two inner rings are beta subunits; these are proteolytically-active • archaeal proteasome • one type of alpha subunit • one type of beta subunit core particle (20S) • the evolution of the proteasome’s subunit complexity therefore parallels that of archaeal/eukaryal prefoldin/chaperonin • archaeal prefoldin, 2 subunit types; archaeal chaperonin, 1-3 types • eukaryotic prefoldin, 6 subunit types; eukaryotic CCT, 8 types • co-evolution with substrates?
Proteasomecomponents 19-3
Proteasome structure N N N N 19-5 • 19S cap (regulatory complex) is only present in eukaryotic proteasome and its crystal structure is unknown • the circled ‘N’ in Fig. (c) and (e) represent the N-termini of archaeal and yeast proteasome • archaea has only alpha and beta subunits whereas eukarya has different homologous alpha and beta subunits active site Proteasomes from eukaryotes and archaea, showing the cap complex (magenta), core complex (blue, where alpha and beta subunits are shown), slice surface (green), active sites (white circles) and N-termini (circled ‘N’s). In (c) and (f), cyan indicates the residues visualized that are closest to the N-termini (threonine 13 and serine 11 respectively). (a) Electron micrograph of proteasome holoenzyme from a representative eukaryote (Xenopus laevis). (b) Medial cut-away view of the Thermoplasma acidophilum proteasome core. The lumen is divided into three chambers, and the central chamber contains the peptidase active sites (red). (c) Ribbon diagram of two Thermoplasma acidophiluma subunits, showing the structure of the pore. (d) Cut-away view of the Saccharomyces cerevisiae proteasome core. (e) Ribbon diagram of two S. cerevisiaea subunits (left: Pre9/Y13; right: Pre10/Prs1). The N-termini of these subunits are shown to occlude the channel. Adapted from Dan Finley, Encylopedia of Life Sciences.
Eukaryotic proteasome catalytic site 19-6 • active sites (shown with white circles) are on three separate beta subunits; threonine residues are critical during catalysis • the proteasome contains three separate proteolytic activities: - trypsin-like (arg, lys) - chymotrypsin-like (tyr, phe) - post-glutamyl (glu) • controversial: the distance between the active site thr residues is 28A, which may determine the length of the proteolytic fragments, i.e., ~ 8 amino acids
11S proteasome regulator 19-7 • proteasome 11S regulator also consist of heptameric rings and bind the 20S core much as the 19S regulatory cap does • also called PA26, PA28 and REG • binding of the 11S particle stimulates proteasomal activity • may facilitate product release by opening proteasome ‘gate’ • reduction in processivity expected for an open conformation of the exit gate may explain the role of 11S regulators in the production of ligands for MHC class I molecules • 11S carboxy-terminal tails provide binding affinity by inserting into pockets on the 20S proteasome, and 11S activation loops induce conformational changes in alpha-subunits that open the gate separating the proteasome interior from the intracellular environment Proteasome co-crystallized with 11S regulator particle Proteasome without 11S regulator Proteasome with 11S regulator smaller peptides larger peptides
Bacterial proteasome-like proteases? 19-8 • MOST Bacteria do not possess proteins that are closely related to the proteasome, but: • HslV is is structurally-related to proteasome • HslU is the regulatory particle • HslVU is responsible for the degradation of the cell division inhibitor SulA; its repertoire of substrates likely includes other cellular proteins HslUV HslU HslV (2 rings) structures of 2 subunits; superimposable to the beta subunits of the archaeal proteasome em picture inside view of HslV protease (active site) • Lon and FtsH have combined regulatory and protease domains into one single polypeptide that assembles into a ring structure • shown to have chaperone-like activity, can disassemble aggregates, and can mediate protein degradation
Bacterial proteasomes - comparison Figure 1 Comparison of different classes of ATP-dependent pro- teases, shown as a side-on cross-section. (A) The eukaryotic 26S proteasome, composed of a 20S core particle (blue a and b subunits) flanked by 19S regulatory subunits (magenta and orange). The 19S subunits bind to the substrate through the covalent ubiquitin modification (yellow) and unfold it by pulling on the unstructured initiation site (bright green). Ubiquitin is removed to be recycled during the degradation process. (B) The bacterial protease ClpXP, composed of rings of the protease ClpP (dark green) and the ATPase motor ClpX (red). ClpX binds to the degradation signal, in this case the ssrA peptide sequence (green), which also serves as the site for the initiation of degradation. (C) The actinobacterial proteasome, consisting of a 20S core particle similar to that of the 26S proteasome, and a single ring of the ATPase Mpa (purple). Mpa binds to the substrate through the covalent Pup modification (light green). Pup has an N-terminal unstructured region, which serves as the site for the initiation of degradation, leading to complete degradation of Pup. (D) Archaeal proteasome is most closely related to the eukaryotic proteasome, with core alpha and beta subunits and Rpt-like AAA ATPase subunits (termed PAN for Proteasome Activating Nucleotidase) involved in protein unfolding. eukaryotic bacterial all bacteria actino-bacteria only D Kraut and Matouschek EMBO J. 2009 archaeal
* *
ClpAP, ClpXP proteases 19-9 • ClpAP and ClpXP are ATP-dependent proteases • ClpA, ClpX are chaperones • ClpP is the protease • substrates: soluble, abnormal proteins • ClpAP and ClpXP can also degrade any protein tagged with SsrA, an 11-residue peptide that is added to arrested chains in bacteria • ClpA and ClpX (in the absence of ClpP) can also disassemble protein complexes (similar to how Hsp104 from yeast can disentangle protein aggregates) ClpAP, ClpXP are ‘active’ proteases ClpP by itself not active as protease - ClpP (2 rings) has been crystallized - ClpA, ClpX attach As single rings on Opposite sides of ClpP • symmetry mismatch • ClpA and ClpX have six subunits per ring; ClpP is a homo-heptameric ring • symmetry mismatch may have implications for activity (but, other proteases don’t have this symmetry mismatch, so relevance is not clear)
ATP-dependent protease mechanisms 19-10 • unfolding, then degradation is a common mechanism to ATP-dependent proteases? • work with ClpAP, ClpXP suggest that this is the case • PAN (Proteasome Activating Nucleotidase) • associated with archaeal proteasome; stimulates its activity • AAA ATPase (as with base of 19S proteasome cap); hexameric ring • work with PAN also suggests unfolding then degradation mechanism • the ATPase subunits of the proteasome regulatory particle • shown to have chaperone activity Braun et al. (1999) Nat. Cell Biol.1, 221-226. • likely also involved in unfolding substrates just before translocation into the core particle next lecture: evidence for ClpA unfolding
Compartmentalization 19-11 • compartmentalization, with respect to protein folding and degradation, refers to the encapsulation of substrates within a cavity, or a shielded environment chaperones • chaperonins possess a cavity that is capped by a cofactor (in the case of GroEL/GroES) or with protrusions (in the case of Group II chaperonins CCT and thermosome) • AAA ATPases are also ring-shaped structures that possess a cavity • prefoldin may partially envelopes substrates • this encapsulation provides shielding of substrate hydrophobic residues • in the case of chaperonins it provides ‘infinite dilution’ for substrates proteases • most oligomeric proteases have a cavity that is shielded fromthe bulk cytosol - e.g., proteasome, HslUV, ClpAP/XP, Tricorn protease, etc. • shielding the active site is necessary to preventunregulated proteolysis • encapsulation may assist processivity of protease folding • newly-made, non-native proteins are shielded from the bulk cytosol by chaperones