Download

1 / 85
870 likes | 1.17k Views
INSTITUTE FOR IMMUNOBIOLOGY. 适应性免疫应答细胞 :T 淋巴细胞 Institute for Immunobiology Department of Immunology Center for Gene Immunization and Vaccine Research Fudan University 洪晓武: 021-54237362 xiaowuhong@fudan.edu.cn. Shanghai, China. T 淋巴细胞. 一、 T 淋巴细胞的表面分子及其作用 二、 T 淋巴细胞亚群 三、 T 淋巴细胞功能.

E N D
INSTITUTE FOR IMMUNOBIOLOGY 适应性免疫应答细胞:T淋巴细胞 Institute for Immunobiology Department of Immunology Center for Gene Immunization and Vaccine Research Fudan University 洪晓武:021-54237362 xiaowuhong@fudan.edu.cn Shanghai, China
T淋巴细胞 一、T淋巴细胞的表面分子及其作用 二、T淋巴细胞亚群 三、T淋巴细胞功能
T细胞是由一群功能不同的异质性淋巴细胞组成,由于它在胸腺内分化成熟故称为T细胞。T细胞是由一群功能不同的异质性淋巴细胞组成,由于它在胸腺内分化成熟故称为T细胞。 成熟T细胞由胸腺迁出,移居于周围淋巴组织中淋巴节的副皮质区和脾白髓小动脉的周围。 T细胞执行特异性细胞免疫应答 并在TD-Ag诱导的体液免疫应答中发挥重要作用
TCR-CD3复合物 TCR可分为TCRαβ和TCRγδ两种类型 结构相似 两条异源二聚体肽链藉二硫键组成的跨膜分子 每条肽链含V、C区,类似Ig结构
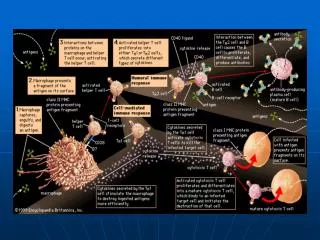
TCR-CD3复合物 是T细胞抗原受体与一组CD3(γGamma δDeltaεEpsilonζZeta ηEta )分子以非共价键结合而形成的复合物,是T细胞识别抗原和转导信号的主要单位。TCR特异识别由MHC分子提呈的抗原肽,CD3分子转导T细胞活化的第一信号
2. CD4和CD8分子 • CD4 主要分布于成熟Th细胞、巨噬细胞、DC细胞等表面; • 是HIV受体,与APC表面MHC-II分子非多态区结合 • CD8 主要分布于成熟Tc细胞表面,与APC表面MHC-I分子 • 非多态区结合 • 既能加强T细胞与APC或靶细胞的相互作用,又能参与抗原刺激TCR-CD3分子信号转导
CD4和CD8 -T细胞辅助受体 加强T细胞与APC或靶细胞的相互作用 通过胞浆区的CxCP基序与p56lck(Src family tyrosine kinase LCK)酪氨酸激酶相连,参与T细胞活化和增殖信号转导
3. 协同(辅助)信号分子 协同信号分子 TCR-CD3复合分子可提供第1信号 协同刺激信号(costimulatouy signal)或第2信号 参与T细胞活化的协同刺激信号主要是CD28--CD80/86,CTLA4--CD80/86给予已活化T细胞抑制信号。
1、CD28和CTLA-4 T APC CD28 CD80/86 CTLA4 CD40L CD40 LFA-1 ICAM-1 LFA-2 LFA-3 (intercellular adhesion molecule-1)
2、ICOS 3、CD40L 4、CD2 5、LFA-1和ICAM-1
4. 其它一些受体 • 丝裂原受体 • 细胞因子受体 • 病毒受体
二、T细胞分化发育 thymic corpuscle
胸腺微环境是诱导并调控T细胞分化发育的关键因素胸腺微环境是诱导并调控T细胞分化发育的关键因素 • 胸腺基质细胞(TSC) • 细胞因子 • 胸腺激素
双阴性期(DN) 原T细胞(pro-T)、前T细胞(pre-T) αβTCR- 、CD3 - 、CD4 - 、CD8 - 双阳性期(DP) αβTCR + 、CD3low、CD4 + 、CD8 + 单阳性期(SP) αβTCR + 、CD3 + 、CD4 + αβTCR + 、CD3 + 、CD8 + 成熟T细胞,具有识别抗原、介导免疫应答及参与免疫调节的功能
1. T细胞发育的阳性选择(positive selection) CD4+CD8+T细胞+胸腺基质细胞(表面MHC分子) 如果与MHC-I结合,最终分化为CD8+T细胞 如果与MHC分子不结合则在胸腺皮质中凋亡 如果与MHC-II结合则最终分化为CD4+T细胞 胸腺细胞经阳性选择赋予成熟T细胞在识别抗原时具有MHC限制性
2. T细胞发育的阴性选择(negative selection) DP或经阳性选择的SP的T细胞 其TCR识别胸腺基质细胞表面高亲和力的MHC或MHC-自身抗原肽的T细胞克隆将发生凋亡 经阴性选择可清除自身反应性T细胞克隆 获中枢耐受
三、T细胞亚群 1)根据TCR种类 αβT、γδT细胞 在末梢血主要为αβT细胞可占95%,而γδT细胞只占1%~10%。αβT细胞为主要参予免疫应答的T细胞,两者特性和功能均不相同。
TCRαβT和TCRγδT细胞 TCRαβT TCRγδT 较少多样性 5-15%,粘膜上皮 成熟大多数CD2CD3 简单多肽、 HSP、脂类、多糖 MHC样分子 Tc TCR 分布 表型 识别抗原 MHC限制 功能 极大多样性 60-70%,外周淋巴组织 成熟CD2CD3CD4/CD8 8-17aa 经典MHC Th、Tc
2)根据T细胞是否表达CD4或CD8分类 CD4+ T细胞或CD8+ T细胞 TCRαβTCD4+细胞:CD2+、CD3+、CD4+、CD8- TCR识别抗原是MHCⅡ类分子限制性 TH0、Th1和Th2、行使Tc、Ts功能 TCRαβTCD8+细胞:CD2+、CD3+、CD4-、CD8+ TCR识别抗原是MHCI类分子限制性 行使Tc、Ts功能
3)功能性亚群:Th、Tc、TDTH、Ts • Th细胞 根据所分泌的细胞因子不同,将其分为Th0、Th1和Th2亚型。 • Tc细胞 杀伤、分泌IFNγ、IL-4、IL-5和IL-10 • Ts细胞 • TDTH 主要为CD4 + Th1
4)初始T细胞和记忆性T细胞 记忆性T细胞表达CD45RO,而初始T细胞表达CD45RA 5)NK1.1 T细胞 其TCR识别的抗原是由CD1分子提呈的脂类和糖脂类抗原 Leukocyte Common antigen
四、T细胞功能 1)免疫调节功能(Th1、 Th2 、 Th3、Ts 、Treg) 2)特异性杀伤功能(CTL、Th1、γδT) 3)介导超敏反应(TDTH) 4)新型效应T细胞:Th17
Multiple subsets of Treg cells Naturally occurring CD4+CD25+ Treg cells (5–6%) GITR and Foxp3 a cell–cell contact mechanism Antigen-induced Tr1 and Th3 cells (IL-10) and/or (TGF-b) ck-dependent mechanism no specific marker has been identified. Adaptively induced CD4+ Treg cells GITR and Foxp3 a cell contact dependent or soluble factor-dependent (other than IL-10and/or TGF-b) mechanism CD8+ Treg cells NKT regulatory T cells glucocorticoid-induced TNFR family related gene (GITR)
Naturally occurring CD4+CD25+Foxp3+ Tregs Development and function of naturally occurring CD4+CD25+ FoxP3+ regulatory T cells (nTregs). Development:bone marrow-derived CD4+ T cell precursors develop naturally into nTregs upon beneficial TCR engagement by self-peptide–MHC complexes and Foxp3 induction in the thymus. Upon instruction in the thymus, nTregs emigrate into the periphery as functionally fully competent cells. Mode of action: upon TCR cross-linking, peripheral nTregs suppress the proliferation and IL-2 production by responder CD25–CD4+ or CD8+ T cells in a contact-dependent manner either (a) directly or (b) indirectly via the APC. In addition: nTregs may (c) condition DC to become tolerogenic and turn down the response of conventional T cells on her part
Extrathymic induction and function of adaptive regulatory T cells. Adaptive regulatory T cells (aTregs) differentiate from naive conventional CD4+ T cells either as a result of suboptimal antigenic stimulation by resting/immature DC, the influence of suppressive cytokines like IL-10, TGF-b or cell contact-dependent interaction with activated nTregs (infectious tolerance). Their mode of action involves both cell contactdependent (Tr1 cells) and contact-independent suppressive activities (Th3 cells). Through the production of IL-10 and TGF-b they convert immature DC into tolerizing APC
Fig. 3 Role of Tregs in early and late stages of microbial infections. In the stages of an immune response against a microbial infection Tregs behave differently. A Throughout the early phase of the response the suppressive activity of Tregs is turned down by effector T cell-derived IL-2 and microbial components such as TLR-ligands. Tregs respond to the stimulation by mature DC and proliferate.
b At the late stage of the response, when the invading organism is cleared from the host, Tregs regain their suppressive function and participate in the silencing of the T cell response by acting on effector T cells and DC. Possibly, this late activity is also for the proper development of memory T cells
Treg-based immune intervention strategies. Selective manipulation of Treg function is an emerging target for immune intervention strategies to either boost responses in cancer and microbial diseases or suppress those unwanted in autoimmunity, allergy, transplantation and pregnancy disorders. The transient depletion of Tregs as well as their modulation by microbial agents may allow a transient reduction of Treg activity and enforce anti-tumor responses and immunity against viral infections. On the other hand their selective activation could diminish chronic pathological immune responses
Generation and conversion of Treg cells in the tumor microenvironment Tumor cells not only provide antigenic stimulation for T cell activation but also interact with tumor-infiltrating innate immune cells to secrete crucial cytokines for T-cell differentiation. Naı¨ve CD4+ T cells can be differentiated into different subsets of CD4+ T cells, including Th1, Th2, Treg and IL-17-producing T cells (Th17), depending upon the strength of antigen stimulation and cytokine milieu. It is known that combination of suboptimal antigen stimulation with TGF-b favors the conversion of naive T cells into Treg cells but blocks the generation of Th1 or Th2 cells. However, TGF-b plus IL-6 facilitate the conversion of naive T cells into Th17 cells. Alternatively, naturally occurring CD4+CD25+ Treg cells directly derived from the thymus can cross-react with some antigens expressed by cancer cells, thus promoting their expansion and accumulation in the tumor microenvironment.
Human IL-17 and IL-17R key features a Two isoforms (long and short).
Therapeutic targets for autoimmune inflammatory diseases are associated preferentially with the IL-23/Th17 pathway
The pathogenic role for IL-23, not IL-12, in mouse models of autoimmunity Studies by Cua and co-workers have demonstrated that disease development requires IL-23, but not IL-12, in EAE and CIA. Compared with wild-type susceptible mice, mice deficient for IL-23 (Il23p19/) and both IL-23 and IL-12 (Il12p40/) failed to develop disease after antigenic challenge, whereas mice deficient for IL-12 (Il12p35/) developed more severe disease.
Model of Th1 versus Th17 lineage development from naive CD4 T cell precursors (Tn) This model emphasizes the distinct lineages leading to mature Th1 and Th17 effector cells (see main body of text for details). Question marks denote speculative or unknown aspects of Th17 differentiation that are yet to be defined.
Antagonistic cytokine networks control CD4 effector T-cell differentiation Recent studies have established that Th1 and Th2 effector cytokines, IFNg and IL-4, respectively, potently inhibit Th17 development. Furthermore, TGF-b, a cytokine previously implicated in Treg development and function, appears to be required for Th17 development, both through indirect effects (blockade of IFNg and IL-4 production by cells of the innate immune system) and through direct effects on naive CD4 T-cell precursors (Tn).
CD4+T细胞分化和免疫调节细胞因子网络模式简图CD4+T细胞分化和免疫调节细胞因子网络模式简图
Diversification of CD4 T Cell Lineages Although functional CD4 T cell development has been dominated by the Th1-Th2 paradigm for nearly two decades, the number of defined lineages has now increased. The cytokines associated with arrows indicate dominant cytokines involved in specification of each of the indicated lineages. The cytokines listed below each cell type indicate key effector or regulatory cytokines produced by differentiated cells of that lineage or, in the case of nTreg, a contact-dependent mechanism of suppression. Tn: naive, postthymic CD4 T cell precursors; Tp: thymic precursors.Dotted lines represent less well-defined lineage relationships.
Model of Branching Th17 and Adaptive Treg Lineage Development This model emphasizes distinct pathways leading to mature Th17 effector cells or Foxp3+ adaptive Tregs (aTreg), induced by a common requirement for TGF-b but differential effects of IL-6 and IL-23. Naive CD4 T cells (Tn) activated by antigen presented on immature DCs that do not produce IL-6 production are induced by TGF-b to express Foxp3 and develop into aTregs (top panel). Tns activated bymature,TLR-activatedDCsthat produceIL-6 are induced by TGF-b to upregulate IL-23R and become competent for IL-17 production and IL-23 signaling. IL-23 signaling induces responsiveness to IL-18 and IL-1, which can act synergistically with IL-23 to induce Th17 cytokineproduction independently ofTCRstimulation.Alternatively, TCR stimulation by antigen can induce Th17 cytokine production directly, without a requirement for IL-23, IL-1, or IL-18.Dotted lines indicate possible positive feedback loops by which cytokine products of Th17 (IL-6) or aTreg cells (TGF-b1) may reinforce lineage development.
CD4+辅助性T细胞 • CD8+杀伤性T细胞
3. 抑制性T细胞 4. 迟发型超敏反应性T细胞(TDTH) 5. NK1.1+T细胞
掌握TCR分型与结构 • 掌握T细胞分群及不同亚群的生物学特性 • 掌握T细胞发育过程 • 熟悉T细胞表面主要膜分子及其作用 • 了解TCRγδT细胞和TCRαβT细胞的异同点
T淋巴细胞对抗原的识别及免疫应答 一、T细胞对抗原的识别 二、T细胞活化的过程 三、效应性T细胞的应答效应
免疫应答的基本过程 抗原识别 抗原受体与抗原的特异性结合 免疫应答 抗原识别、反应、效应的全过程 免疫反应 免疫效应物质与抗原结合的过程
一、T细胞对抗原的识别 T 细胞抗原受体及其识别抗原的特点 • 只识别表达于APC表面并与MHC分子结合 成复合物的多肽 • 只识别氨基酸一级序列的多肽线性决定簇 • TCR识别抗原受到MHC的限制—— CD4+T只识别与MHC-II分子结合的肽段 CD8+T只识别与MHC-I分子结合的肽段