Deciphering Oxidative Phosphorylation Journey Through History
210 likes | 280 Views
Explore the history, intricacies, and mysteries of oxidative phosphorylation and photophosphorylation processes in biochemistry. Learn about electron transferring, ATP biosynthesis, coupling of reactions, and the evolution of scientific understanding over the years. Delve into the molecular processes and key discoveries that have shaped our current knowledge in this fascinating field of study.

Deciphering Oxidative Phosphorylation Journey Through History
E N D
Presentation Transcript
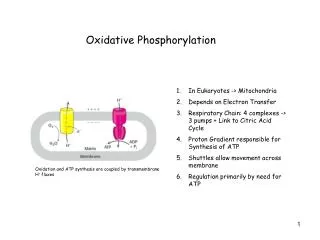
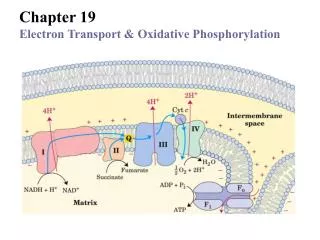
For Biochemistry II lectures of Nov. 26 and Dec. 3, 2008 (Prof. Zengyi Chang) Chapter 19 Oxidative Phosphorylation Electron transferring (flow ) through a chain of membrane bound carriers (coupled redox reactions),generation of a transmembrane proton gradient,ATP biosynthesis (ADP phosphorylation). “Anyone who is not confused about oxidative phosphorylation just doesn’t understand the situation” Efraim Racker, 1970s
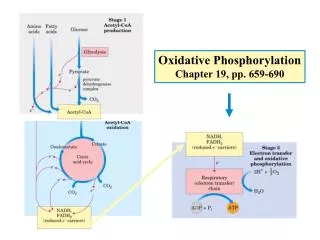
The three stages of biological oxidation Main events: *all the electrons are transferred to O2; *ATP is made using a proton gradient. Energy-releasing Energy-requiring Coupling of energy-releasing & energy-requiring reactions.
History of understanding oxidative phosphoryation • 1900s: vital role of phosphate in fermentation revealed. • 1910-1920S: role ofironin intracellular respiration realized. • 1920s: Cytochromesidentified and the concept of therespiratory chain (electron transport chain)formulated. • 1930s: pyruvate (product of glycolysis) known to be completely oxidized to CO2 via thecitric acid cycle(needing O2). • 1930s: NAD+andFADwere found to be e- carriers between metabolites and the respiratory chain. • 1930s: role ofATPand general importance ofphosphorylationin bioenergetics realized (Lipmann,1939, “oxidative phosphorylation” introduced)
A historical perspective • Understanding the detail molecular process • 1940s: link between sugar oxidation and ATP synthesis established; Role of NADH linking metabolic pathway and ATP synthesis proved. • 1950s: isolatedmitochondriafound to effect theobligatory couplingof thephosphorylation of ADPandthe e- transfer from NADH to O2. • 1940s-1950s:“High energy intermediate” leading to ATP synthesis (chemical coupling hypothesis) was searched (but failed to be found.) • 1960s: thechemiosmotic hypothesisproposed for linking the e- transfer and ADP phosphorylation (role of the membrane and the across-membrane proton gradient postulated, a new paradigm!). • 1970s: thebinding change modelwas proposed to explain how the proton gradient will be used to drives ADP phosphorylation. • 1990s:structure ofATP synthasedetermined, supporting the binding changemodel.
Electrons of NADH and FADH2 are transferred to O2 via many intermediate electron carriers making up the respiratory chain.
History of understanding Photophosphoryation • 1790s: CO2 and H2O are taken up by green plants, while O2 is released, all under the influence of light. • 1930s: photosynthesis occur via light-dependent redox reaction; isolated chloroplasts made O2 but did not fix CO2; O2 is derived from H2O. • 1950s: The path of carbon fixation revealed. • 1960s: Existence of two photosystems elucidated.
NADH enters at NADH-CoQ oxidoreductase Consisting ~34-46 polypeptide chains ~880 KDa(the largest among four) NADH: 2e FAD:1e FMN 1e or 2 e The coupling mechanism between e- transferring and H+ pumping is still unknown!
4Fe-4S 2Fe-2S At least eight different types of iron-sulfur centers (first revealed by Helmut Beinert) act in the respiratory chain: (in complex I, II and III) iron atoms cycle between Fe2+ (reduced) and Fe3+ (oxidized).
The isoprenoid tail Ubiquinone (or coenzyme Q)is the only e- carrier that is not bound to a protein and is able to diffuse freely in the lipid bilayer . Q10 (or dihydroubiquinone)
FADH2 of flavoproteins also transfer their electrons to ubiquinone (Q), but with no H+ pumped. Succinate dehydrogenase
Structure of mitochondrial complex II (porcine) was determined Succinate dehydrogenase Sun F et al. and Rao, Z, 2005, Cell, 121:1043-1057. 孙飞 Proposed electron path
The crystal structure of the cytochrome bc1 complex (Complex III) has been determined. Structure of the three core subunits out of the 11 subunits. Spatial relationship of the cofactors deduced from the resolved structure of complex III. Science, 1997, 277:60-66.
Three types of heme groups are found in the cytochromes. Also found in Hemoglobin & myoglobin Bound to cytochromes tightly but noncovalently Bound to cytochromes tightly and covalently Bound to cytochromes tightly but noncovalently The iron interconverts between its reduced (Fe2+) and oxidized (Fe3+) froms, thus performing oxidation and reduction reactions
Reduced cytochromes has three absorptionbands in the visible wavelengths Cytochromes are classified on the basis of position of their lowest energy absorption band in the reduced state. c b a The reduced (Fe2+) state of cytochromes a, b, and c has the longest wavelength band near 600, 560, and 550 nm respectively.
The e- transferring & H+ pumpingin Complex III occur via the Q cycle For each 2 e- transferred, 4H+ are translocated.
the Q cycle From: http://en.wikipedia.org/wiki/Image:Theqcycle.gif
The three critical subunits (out of 13) of complex IV. Cytochrome c oxidase (Complex IV ), contains 3 Cu and 2 heme A groups as electron carriers. CuA -CuA a CuB a3 Heme a and heme a3 has identical structures but different reduction potential. O2 acts as the final electron acceptor here. Science, 1996,272:1136-1144
Four electrons are transferredfrom 4 Cyt c to 1 O2 to make 2H2O in Complex IV,with 4H+ taken from the matrixto make 2H2O&4H+ pumped out.
~ 10 protons per NADH and ~6protons per FADH2 oxidizedare pumped across the inner membrane of mitochondria (or plasma membrane of bacteria) The highly mobile Q and Cyt c molecules shuttle electrons from one large multiprotein complex to another. The electron motive force is converted to an proton motive force. A transmembrane H+ gradient is thus generated.