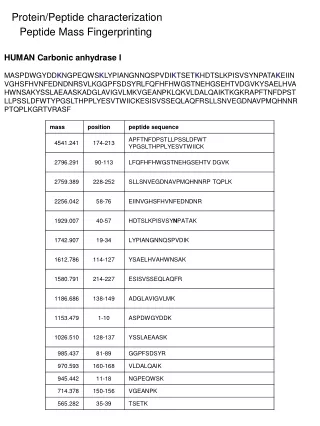
Peptide semnal
Peptide semnal Atunci când o proteină este sintetizată, structura sa secundară sau terţiară se construieşte din nou şi pe măsura transducţiei este eliberată în citoplasmă.


Peptide semnal
E N D
Presentation Transcript
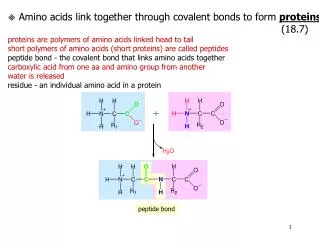
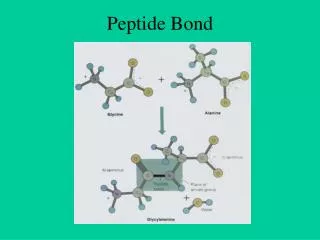
Peptide semnal Atunci când o proteină este sintetizată, structura sa secundară sau terţiară se construieşte din nou şi pe măsura transducţiei este eliberată în citoplasmă. Dacă această proteină este destinată pentru a fi în membrane sau organite celulare, partea codantă a ARNm începe printr-o secvenţă de câţiva aminoacizi ca adresă pentru încorporarea acestei proteine în membrane. Peptidele semnal sunt lanţuri formate din câţiva aminoacizi situaţi la extremitatea NH2 terminală a proteinelor traduse, semnalând unde trebuie să fie excretată proteina din celulă.
Aceste peptide orientează destinaţia proteinelor; încorporate în membrană (RE, AG, plasmalemă, lizozomi) intră în mitocondrii sau sunt exocitate în afara celulelor prin aparatul Golgi. Pentru că funcţia lor este de a penetra prin membrana RE structura acestora este bogată în aminoacizi hidrofobi. • Fiecare organit prezintă un mecanism de recunoaştere şi încorporare numai a proteinelor necesare şi specifice lui.
Această recunoaştere se realizează datorită unei secvenţe de aminoacizi pe care o conţine proteina respectivă, • secvenţa semnal, şi a unui receptor specific care recunoaşte proteina şi o internalizează. • După ce proteina a ajuns la destinaţie, secvenţa semnal este eliminată prin digestie proteolitică, iar în cazul receptorului, acesta este eliberat şi reciclat.
Poliribozomii şi peptidele semnal • Poliribozomii care se formează pe un mesager conţinând o peptidă semnal, au o evoluţie uşor diferită. • Transducţia se opreşte cu puţin după sinteza peptidei semnal. • Acest lucru este posibil datorită particulei de recunoaştere a semnalului SRP (Signal Recognition Particle)
. SRP este constituit din diferite proteine şi un ARN 7S. Imediat ce interacţiunea peptidei semnal cu SRP apare în afara structurii ribozomilor, şi se leagă cu SRP este inhibată elongaţia. • Traducerea se opreşte până când SRP este recunoscut de un receptor al membranei RE. • Imediat ce această legătură este stabilită ribozomul este legat de ribozomii din membrana RE lângă un complex proteic. • Acesta contribuie la formarea unui canal apos de transport al proteinelor cunoscut sub numele de translocon, prin care proteina nou formată trece în lumenul, apoi în cisternele RE.
Deci canalul se deschide şi elongaţia se reia. • La faţa internă a membranei RE o endopeptidază specifică sau peptidaza semnal va secţiona peptida semnal şi sinteza se va desfăşura până la terminare. • SRP se eliberează în citosol şi traducerea este încheiată. • Proteina va fi în final încorporată într-o membrană sau exportată pentru traversarea AG către exteriorul celulei.
Adresarea proteinelor către mitocondrii face apel la un alt tip de peptidă de adresare care conduce peptidele prin traversul membranei mitocondriale. • Produşii primari de traducere conţinând peptide semnal sunt denumiţi preproteine, de exemplu preproinsulina, hormonul preproparotidian, etc.
Organizarea structurală a proteinelor • Lanţurile polipeptidice ale proteinelor se pliază în diferite moduri atât în cadrul propriului lanţ cât şi între lanţurile vecine adică intracatenar şi intercatenar. • Mult timp s-a considerat că modurile de pliere ale lanţurilor polipeptidice sunt determinate numai la secvenţa aminoacizilor din lanţ. • Astfel s-a stabilit că proteinele având aceeaşi secvenţă a aminoacizilor pot exista în forme diferite de împachetare şi că astfel de plieri pot fi influenţate de prezenţa altor proteine cunoscute sub numele de molecule supraveghetoare tip scufiţă (chaperones).
Denaturarea şi renaturarea proteinelor • O proteină care posedă o proprietate biologică proprie, unică, se numeşte proteină nativă şi ea se deosebeşte de proteina ce şi-a pierdut această proprietate şi pe care o numim de aceea denaturată. • O proteină denaturată şi-a pierdut structura tridimensională sau în alţi termeni conformaţia sa. • Denaturarea poate fi reversibilă sau ireversibilă.
Denaturarea reversibilă se poate realiza prin folosirea atentă a unor reactivi ca ureea şi mercaptoetalonul. • Ureea distruge structura apei şi de aceea limitează interacţiunile hidrofobe ale catenelor laterale R şi a resturilor de aminoacizi, ceea ce duce la deplierea şi la disocierea moleculelor proteice. • Mercaptoetanolul reduce legăturile S-S. De aceea la îndepărtarea ureei şi a mercatoetanolului, proteina s-ar putea renatura. • Renaturarea este interpretată ca o dovadă în sprijinul ipotezei că o proteină având o structură primară corectă se va plia în mod spontan conducând la structura unică responsabilă pentru activitatea sa biologică.
Astăzi se ştie că renaturarea proteinelor poate fi asistată pe două căi. • Una implică proteina disulfid izomeraza o enzimă care are rolul de a corecta legăturile S-S greşit formate. • A doua cale implică structurile tip chaperone. • Ele se pot defini ca o familie de proteine din clase neânrudite care mediază asamblarea corectă a altor polipeptide dar care nu sunt componente ale structurilor funcţionale asamblate.
Ciclul celular • Toate celulele vii urmează fie un un program de diviziune, fie o moarte programată, numită apoptoza. • interfaza perioadă în care celulele cresc şi mitoza, perioada în care nucleul şi restul celulei se divide. • Viaţa celulei se derulează între 2 mitoze. La mamifere această perioadă durează mai puţin de 30 de ore; sunt celule a căror viaţă este foarte scurtă sau foarte lungă.
Pe durata acestor 30 ore, celulele traversează 4 faze: • faza G1 unde genomul fiind diploid, fiecare genă este reprezentată în 2 exemplare. Cromatina este accesibilă ARN polimerazelor. • Faza S la jumătatea ciclului începe replicarea ADN; ADN –polimeraza va acţiona în jur de 8 ore pentru a recopia în dublură ADN fiecărui cromozom. • Masa celulară creşte continuu în timp ce conţinutul de ADN creşte în faza S şi scade brusc după faza S, astfel că ADN din celulele care nu se divid este constant. • celula intră în faza G2 unde fiecare genă este reprezentată în 4 exemplare.
Cromatina este din nou accesibilă ARN polimerazei care reâncepe transcripţia; • survine mitoza care dă naştere la două celule fiice. Fiecare va primi una din copiile identice din ADN fiecărui cromozom şi fiecare genă va fi reprezentată în două exemplare. • Ciclul celular are trei puncte de decizie: la punctul G1, G2 şi la sfârşitul mitozei • Proteina Cdk (proteinkinaza dependentă de cicline) a fost identificată ca principalul reglator al trecerii prin aceste trei puncte.
În G1există unul din principalele puncte de control care decide dacă celula ar trebui să se dividă sau rămâne în acest stadiu. • Complexul Cdk prin intermediul kinazelor fosforilează şi activează numai anumiţi factori de transcripţie pentru cicline. • Nivelul ciclinelor în G1 creşte şi sunt asociate cu Cdk complex numit şi CdkG1. • La începutul fazei G1 complexul de prereplicare ADN este defosforilat de Cdk G1 şi se asamblează în originea de replicare a cromozomului. • O ţintă a acestor factori de transcripţie sunt genele care codifică componentele Cdk pentru faza S.
În acest punct funcţia Cdk-S este blocată de inhibitori specifici.Startul pentru faza S este dat de fosforilarea inhibitorilor pentru ciclinele Cdk G1. • Triplarea nivelului Cdk G1 determină trecerea celulei în faza S, de replicare a ADN. • Complexul Cdk-S fosforilează complexul de prereplicare inducând iniţierea replicării. • Fiecare cromozom este replicat, formând 2 cromatide surori conectate prin molecule coezive.
Complexul Cdk M mitotic se formează în timpul fazei S şi a fazei G2 dar activitatea lui este inhibată până când sinteza ADN este completă. • -În G2 este al doilea punct de control. Se apreciază modul în care s-a desfăşurat replicarea ADN şi în funcţie de aceasta se dă startul mitozei. Dacă acest punct este depăşit începe mitoza. • Principala funcţie al MPF –factorul de fosforilare mitotic - este de activare a proteinelor care distrug ciclinele. • În timp ce mitoza se derulează, la sfârşitul metafazei nivelul Cdk este relativ constant dar ciclinele G2 sunt degradate cauzând scăderea lor progresivă, MPF devine disponibil şi iniţiază mitoza. • După mitoză, acumularea treptată a noilor cicline dau startul unui nou ciclu celular.
-Al treilea punct este atins la sfârşitul metafazei. Celula trece astfel de la mitoză la interkineză în stadiul G1. • Activitatea Cdk M începe în timpul mitozei şi constă în fosforilarea mai multor substraturi. • -Complexul Promotor al Anafazei – CPA- este activat şi degradează coezina reglatoare urmată de segregarea cromatidelor surori. • Acest complex degradează de asemenea ciclinele din complexul Cdk în finalul mitozei.
Rolul factorilor de creştere în diviziunea celulară • Creşterea celulară este determinată de legarea proteinelor reglatoare numite factori de creştere care stimulează diviziunea celulară. • Celula prezintă la suprafaţă receptori specifici care recunosc factorii de creştere. • Factorul de creştere dă startul sistemelor semnal intracelulare. • Legătura factorului de creştere provoacă o mişcare în cascadă a semnalelor intracelulare, determinând fosforilarea proteinelor care activează proteinele reglatoare nucleare şi se declanşează diviziunea celulară.
Cromatina şi ADN • În cursul fazelor ciclului celular, cromozomii evoluează pentru a pregăti mitoza. • În timpul G1 cromatina este descompactată şi genele se pot exprima. • Fiecare cromozom nu conţine decât o cromatidă. • În timpul S buclele de replicare se deschid şi începe replicarea. • Sunt două cromatide pentru un cromozom. În timpul mitozei centromerul se leagă de fusul acromatidic şi pregăteşte separarea. • Cromatina este compactată la maxim; după mitoză, cromatina este decompactată şi genele se pot exprima în fiecare din celulele fiice.
REPLICAREA • ADN se replică în celulă printr-un proces care asigură ca una din catenele parentale să fie prezentă în celulele fiice, aşa numita replicare semiconservativă. • În cursul vieţii celulei (ciclul celular, diviziunea mitotică următoare) ADN trebuie să fie dedublat pentru că fiecare celulă fiică să capete un genom complet în nucleul său.
Această sinteză se produce în faza S (în mijlocul ciclului celular) ca urmare a activităţii ADN polimeraza şi . • Replicarea semiconservativă • Cele 2 lanţuri de ADN parental în curs de replicare servesc fiecare ca model pentru sinteza noului lanţ. • În acest mod 2 catene în loc să rămână ansamblate la fiecare sinteză (replicarea conservativă) se separă totdeauna la fiecare ciclu (replicare semiconservativă. • La prima generaţie o catenă a fiecărui dublu helix provine din celula mamă. La a II-a generaţie nu există mai mult de două catene ADN a celulei mame, 4 dublu helixuri nou formate.
Enzimele implicate în replicare sunt: • ADNpolimeraze, topoizomeraza, helicaza, ARN primaza, ADN ligaza. • ADN este sintetizat de enzime numite ADN polimeraze, fiecare dintre acestea utilizând dezoxinucleozid trifosfaţi ca substraturi; polinucleotidul este sintetizat în direcţia 5'-3'. • Matriţa de ADN este utilizată pentru a direcţiona ordinea bazelor azotate în polinucleotidul nou sintetizat care devine complementar cu ADN parental.
Topoizomeraza I şi helicaza • Replicarea ADN necesită o fuziune parţială în dublu helix care modifică înrularea celor 2 catene. • Pentru a permite această reacţie, topoizomeraza este capabilă să modifice rularea hidrolizând o catenă de ADN şi reconstituind-o după ce a fost derăsucită local. • În cursul replicării şi al transducţiei pasul helixului diminuează. • Helicază catalizează dezrăsucirea lanţului dublu catenar, dependent de energia ATP.
Primaza şiADN polimeraza • Ambele enzime au un rol important în replicare la eucariote. • Demararea acţiunii primazei are loc la extremitatea 3OH terminală a lanţului ADN • Ultima ribonucleotidă a amorsei, legată de catena ADN care serveşte ca model, va servi ca punct de iniţiere a activităţii ADN polimeraza . • Amorsa este creată pornind de la o polimerază ARN paticulară (fără legătură cu cele de transcripţie) sau primaza, care poate începe prin a sintetiza 10 nucleotide a unui ARN hibrid, folosind un ADN model. • Prima nucleotidă a acestei amorse se păstrează, cei 3 fosfaţi se prind la capătul celei de-a10-a ribonucleotide a ADN polimeraza şi începe condensarea a 20 dezoxiribonucleotide.
Amorsa va fi construită deci dintr-o catenă mixtă ARN – ADN, din circa 30 nucleotide. • ADN ligazele • ADN ligazele sunt enzime care sunt capabile de reconstituirea legăturilor fosfodiester între 3OH şi fosfatul 5 a două nucleotide vecine dintr-un lanţ de ADN. • Ele intervin în replicare pentru a lega ansamblul lanţului ADN sau fragmentelor Okazaki sintetizate de ADN polimeraze. • Intervin de asemenea în numeroase procese de repararea a ADN genomic.
Se cunosc trei tipuri de ADN polimeraze la E. coli. • ADN polimeraza III joacă rolul major în sinteză, • ADN polimeraza I, lizează amorsa şi completeaza această zonă cu dezoxiribonucleotide. • ADN ligazajoacă un rol atât în sinteza in vivo a ADN, cât şi în repararea unor rupturi monocatenare ale ADN. • ADN procariotelor şi ADN mit prezintă o singură origine a replicării.
Sistemul multienzimatic pentru sinteza ADN • Cel puţin 15 enzime şi proteine acţionează în furca de replicare bidirecţională a ADN la E.coli corespunzând la peste 30 de polipeptide. • Proteina dnaA se leagă la ADN după care se ataşează la un complex al proteinelor dnaB şi dnaC. • dnaB este o helicază care catalizează dezrăsucirea lanţului dublu catenar, dependent de energia ATP. Porţiunea monocatenară liberă a ADN este apoi stabilizată de proteinele de legare SSB (Single Stranded Binding protein). • O amorsă ARN este sintetizată de către primază într-un ansamblu multisubunitar numit primozom. La terminarea replicării amorsa va fi eliminată de către ADN prin polimeraza I.
Un nou ADN este sintetizat de către polimeraza III, holoenzimă formată din opt lanţuri polipeptidice. • ADN aflat în faţa polimerazei pe catena conducătoare nu este afectat de helicază (proteina Rep). • Pe lanţul întârziat, spaţiile lipsă dintre fragmentele de ADN sunt umplute de ADNpolimeraza I şi fragmentele sunt unite de ADNligaze.
Replicarea ADN dublu catenar la eukariote • Pentru a explica creşterea aparentă a ambelor catene a ADN în aceeaşi direcţie în timp ce enzimele sintetizează noi catene numai într-o direcţie 5'-3', s-a postulat că una dintre catene a fost întâi sintetizată în fragmente. • Când ADN a fost examinat la scurt timp după startul sitezei, au fost găsite mici fragmente numite Okazaki după numele celui care le-a descoperit.
Rolul ARN în biosinteza ADN • Sinteza oricărei molecule de ADN este iniţiată prin sinteza unui lanţ scurt de ARN, din nou în direcţia 5'-3', folosind nucleozid trifosfaţi (NTPs) ca substrat, prin intermediul unei enzime numită primaza. • Enzima este selectivă privind situsul de iniţiere a sintezei. • În catena conducătoare numai o amorsă este sintetizată. • În catena întârziată, sunt implicate multe locuri de iniţiere pe măsură ce catenele de ADN parental se separă.
Amorsa ARN este eliminată de ARN polimeraza I şi spaţiile lipsă sunt completate prin acţiunea aceleeaşi enzime. • Fragmentele sunt unite de ADN ligază. • Acest mecanism duce la probleme legate de replicarea capetelor cromozomiale, numite telomere. • Telomeraza rezolvă această problemă.
Bucla de replicare la eukariote • ADN polimerazele încep sinteza în numeroase puncte de iniţiere. • În urma legării proteinelor specifice, dublul helix se deschide pentru a permite demarajul. • Sinteza ADN începe pe amorsa ADN/ARN constituită de primază şi ADN polimeraza. • Replicarea se desfăşoară într-o direcţie – în acest sens unul din cele 2 lanţuri de ADN (lanţul sens) este parcurs de enzimă în sensul 3-5 (pe catena antisens), ceea ce permite sinteza unui alt lanţ în direcţia 5-3şi catena se numeşte lider, leading. • ADN ligazele asigură apoi legarea între diferitele fragmente ale ADN nou.
Sinteza celeilalte catene (lanţul întârziat catena lagging) este mult mai complexă deoarece enzima parcurge acest lanţ de la 5-3. • Primaza şi ADN polimeraza sintetizează o amorsă de 30 nucletoide înainte de zona de replicare şi ADN polimeraza construieşte secvenţe mici de fragmente de ADN în sensul 5-3 (în jur de 200 nucleotide- fragmentele Okazaki). • Ribonucleazele distrug amorsele ARN (fragmentele Okazaki sunt unite între ele prin ADN ligaze). • Există 10 izomeraze ale ADN polimerazei la eucariote. ADN polimeraza asociată polimerazei , este principala enzimă a replicării care sintetizează pe catena directă dar şi pe lanţul întârziat.
Cel puţin 4 ADN polimeraze au fost identificate la eucariote. • Acestea sunt denumite prin litere greceşti α şi β sunt implicate în sinteza ADN nuclear, β în reparare şi γ în replicarea ADN mitocondrial. • Dacă genomul unui virus este format din ADN, replicarea are loc în mod similar cu cel descris la o celulă normală. • Virusurile au capacitatea de a dirija şi utiliza metabolismul normal al unei celule astfel încât în cazul fagilor de liză, de exemplu, infecţia cauzează înlocuirea replicării ADN cu replicarea ADN fagic.
Furca de replicare • Replicarea începe prin separarea celor două catene ale ADN prin helicaze. • Fiecare din cele două lanţuri sunt stabilizate prin SSB. • Pe catena directă, urcând de la 3 la 5, o ADN polimeraza şi o primază sintetizează un lanţ complementar adăugând dezoxiribonucleotide trifosfat la extremitatea 3OH liberă. • Un nou dublu helix se formează între lanţul matriţă directă şi noua catenă sintetizată.
Pe lanţul întârziat o polimerază, ADN polimeraza şi o primază progresează de la 53’ Pentru a putea sintetiza un lanţ complementar, trebuie ca primaza şi ADN polimerazei , să fabrice amorse destul de apropiate la câteva sute de nucleotide distanţa pe lanţul matriţă. • Începând de la 3OH a unei amorse ADN polimeraza sintetizează un fragment Okazaki până când întâlneşte extremitatea 5 trifosfat a amorsei precedente.
Amorsa este hidrolizată de o nuclează ceea ce permite ADN polimerazei să realizeze sinteza unui fragment Okazaki, pe care ADN ligaza îl va lega definitiv pe ADN în fragmentul definitiv. • Un nou dublu helix se formează între catena matriţă directă şi noul lanţ sintetizat. • Toate aceste proteine sunt asociate într-o structură a nucleului (uzina de replicare) care traversează moleculele de ADN. • Această uzină de replicare conţine enzime de preparare a substratului, nucleotid kinaze, ribonucleaze, reductaze etc.







![Peptide [ M]](https://cdn2.slideserve.com/4603643/slide1-dt.jpg)