Download

1 / 63
630 likes | 765 Views
For Biological Macromolecules :. For Biological Macromolecules : Motion is an integral part of function. For Biological Macromolecules : Motion is an integral part of function Motion is good for theoreticians like me. For Biological Macromolecules :

E N D
For Biological Macromolecules: • Motion is an integral part of function
For Biological Macromolecules: • Motion is an integral part of function • Motion is good for theoreticians like me
For Biological Macromolecules: • Motion is an integral part of function • Motion is good for theoreticians like me • Motion is always bad for experimental • structural biologists
Challenges: • Motions occur over a wide range of length scale,
Challenges: • Motions occur over a wide range of length scale, • Structural data are available at varying resolutions,
Challenges: • Motions occur over a wide range of length scale, • Structural data are available at varying resolutions, • How do we simulate, refine & model structures?
Simulating, Refining & Modeling Supermolecular Complexes at Multi-resolution and Multi-length Scales Jianpeng Ma Baylor College of Medicine Rice University
I. Simulation and Refinement at Multi-resolution Scales Quantized Elastic Deformational Model (QEDM) Proc. Natl. Acad. Sci. USA 99:8620-5 (2002) modeling structural motions without atomic coordinates and amino-acid sequence
Procedures of QEDM • Discretize low-resolution density maps by • Vector Quantization or • Cubic grid points of cryo-EM density maps Apply elastic normal mode analysis to the discretized density maps. For very low-frequency deformational modes, the number of points can be significantly smaller than the number of amino-acids.
5 Å 7 Å 15 Å B-factors
Atomic Displacement of Low-frequency mode Standard NMA QEDM at 5 Å QEDM at 7 Å QEDM at 15 Å
Pyruvate Dehydrogenase Complexes (25Å) Truncated E2 core Zhou et al, J. Biol. Chem. 276, 21704-21713 (2001).
PDC is an extraordinarily flexible system Conformational distribution of PDC complex from cryo-EM Zhou et al, J. Biol. Chem. 276, 21704-21713 (2001).
Human Fatty Acid Synthase (FAS) at 19 Å Resolution Proc. Natl. Acad. Sci. USA 99:138-43 (2002)
Experimental Verification & QEDM-assisted cryo-EM Refinement
Conclusions of QEDM: • Capable of simulating low-frequency deformational motions of proteins based on low-resolution density maps. • Provide useful insights into protein functions in the absence of detailed atomic model. • Provide a means to aid structural refinement in cryo-EM measurements.
II. Simulation and Refinement at Multi-length Scales Substructure Synthesis Method (SSM) Proc. Natl. Acad. Sci. USA 100:104-9 (2003) modeling structural motions of filamentous systems from angstroms to microns
Modal Synthesis Procedure in SSM • Compute substructure modes by standard normal mode analysis. • Substructures are assembled by imposing geometric boundary conditions. • Calculate the modes for assembled structure by Rayleigh-Ritz principle. • Focus on a set of low-frequency modes. • Does not need to compute Hessian matrix for the assembled structure.
G-actin monomer A 13-subunit repeat of F-actin filament 37.5 Å
Selected boundary points across the interface filament filament
Lowest-frequency modes in the synthesized system Bending Twisting Stretching
Refining Fibre Diffraction Data by Long-range Normal Modes
In Traditional Fibre Diffraction Refinement: • The filaments are assumed to be a straight helix. • But the filaments like F-actin or DNA molecules deform due to their high flexibility.
Challenge: How do we find proper structural parameters to model the filamentous deformations without overfitting the data?
We chose long-range normal modes of the filaments as refinement parameters.
G-actin monomer A 13-subunit repeat of F-actin filament 37.5 Å
Lowest-frequency modes in the synthesized system Bending Twisting Stretching
Refinement based on long-range normal modes Helical selection rule: l=tn+um t=6, u=13 (conventional method) t=6 (or 12, …), u=1 (our method) l: layerline index n: order of Bessel functions m: any integer t: number of helical turns u: number of asymmetric unit in one crossover
Refinement by single low-frequency vibrational normal mode (13-subunit repeat normal modes)
Conclusion: • Normal modes are good collective variables as structural parameters for refinement. No overfitting of data!!! • Bending motions dominate the contributions, i.e. the filament wiggling motions must be included in the refinement and errors from them can not be compensated from adjusting other local structural parameters.
III. Refinement of Anisotropic Temperature Factors for Supermolecular Complexes in x-ray Crystallography
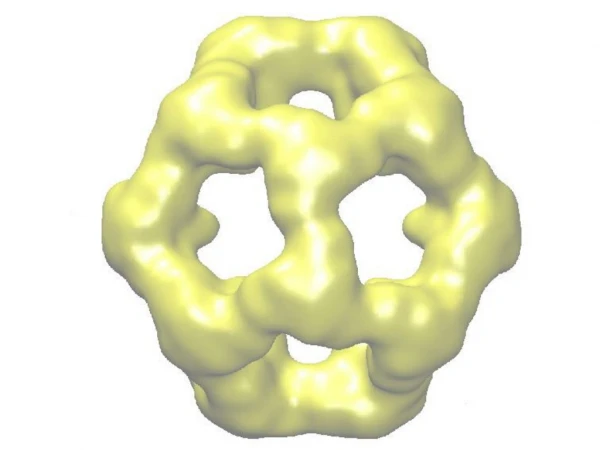
Molecular Chaperonin GroEL GroES GroEL 3 175,000 A 3 85,000 A