Decoding Genetic Information Flows at RNA Level
540 likes | 586 Views
Dive into the intricate world of mitochondrial gene expression, uncovering recoding processes at the RNA level in Diplonema species. Explore unconventional gene structures and evolutionary adaptations in these micro-eukaryotes.

Decoding Genetic Information Flows at RNA Level
E N D
Presentation Transcript
Massive gene defects compensated at the RNA levelGertraud BurgerUniversité de MontréalRobert-Cedergren Centre for Bioinformatics and GenomicsMontreal, QC, Canada
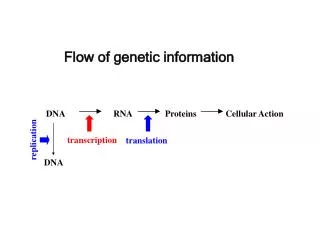
Genetic information flow Central dogma of molecular biology: DNA to RNA to Protein Francis CRICK’s unpublished 1956 sketch of the central dogma. (Image: Welcome Library, London). Cobb 2015 “Who discovered mRNA” Curr Biol 25:R526
Genetic information flow ctn’d genomic To investigate and organism’s makeup we sequence genomic DNAto infer genes& conceptually translate proteins; BUT, certain genes are unrecognizable or defy inference of protein sequence due to information recoding. m Today’s topic: recoding at RNA level in mitochondria.
Overview of talk 1. What is known about mitochondria • genomes • gene expression • intron splicing 2. Unusual mitochondrial genome & expression in a micro-eukaryote: • diplonemids • broken & mutated genes • compensation of defects 3. Evolutionary trends. Mitochondrion Eukaryotic cell
Mitochondrion-encoded genes Intro: mt-genomes ~1,000 proteins in total, <10% mtDNA-encoded Electron transport ATP synthesis Protein synthesis • RNA processing Protein import, assembly, maturation Transcription. Orange, always mtDNA-encoded; Pink, variably mtDNA-encoded. Blue, nucDNA-encoded. Review: Burger et al 2003 Trends Genet 19
Intro: mt-genomes Diversity of mtDNAs Number of distinct chromosomes • 1 (1 to >100) Chromosome copy number • ~10(10 to ~1000) Size • 50 kbp (6 - 11,300 kbp) Shape • circular-mapping linear-tandem • (truly circular; linear monomeric) Gene number • ~35 (5 to 100) • Introns • variable (0 to ~50; Group I, II). Review: Lang et al 1999 Annu Rev Genet 33 More about introns…
Intro: mt gene expression Introns – splicing – cis & trans • Genes can contain introns cis-splicing • Genes can be split within introns trans-splicing • cis-and trans-splicing - same mechanism and machinery. Contiguous gene Split gene e1 e2 ◄ DNA ► ◄ precursor RNA ► ◄ mRNA ► conventional (cis-) splicing conventional trans-splicing e1 e2
Intro: mt gene expression Intron types ...exonGT..intron..AGexon… Spliceosomal introns (nucleus) ‘Archaeal’ (tRNA) introns (nucleus) Group I introns (organelles) Group II introns (organelles). ‘Archaeal’ Group I Group II 3ʹ 5ʹ
Talk’s roadmap √1. What is known about mitochondria • genomes • gene expression • intron splicing 2. Unusual mitochondrial genome & expression in a micro-eukaryote: • diplonemids • broken genes & repair • mutated genes & repair 3. Evolutionary trends. The organism…
Taxonomic group: diplonemids Intro: diplonemids • Morphology: • Flagellated micro-eukaryote, ~10 μm long; • Feeding strategy: • probably scavenging • Ecology: • ocean, any depth • Taxonomy: • previously only 3 genera. Courtesy of B. Leander Highly prolific as we know today…
Every 6th planktonic eukarote is a diplonemid Diplone-mids One of the most abundant and most species-rich marine eukaryotes. de Vargas et al 2015, Science
Diplone-mids Diplonemids in eukaryotic tree Classification: Discoba Euglenozoa Diplonemids Euglenozoa You are here Keeling et al 2005, Trends EcolEvol20 Our research…
Our research • Aim • Explore novel types of gene structure and expression • System • mitochondria of diplonemid type species Diplonema papillatum • Experimental approach • nuclear and mitochondrial genome & transcriptome sequencing • Northern hybridization, RNAase T1-mapping • bioinformatics sequence analysis. Diplonema papillatum Diplonema ambulator Rhynchopus euleides Sulcionemaspecki Diplonemids Lacrimialanifica Flectonemaneradi Hemistasia phaeocysticola
Most unusual mtDNA in Diplonema Instead of an orthodox single 15-50 kbp chromosome: ~100 distinct chromosomes of two size classes: 6 kbp (A), 7 kbp (B) Only ~0.5 kbp unique sequence per chromosome, otherwise repeats WHERE ARE THE GENES ? A B Maslov & Simpson 1999, Protist 150 ; Marande et al 2005, EukaryotCell 4
Our research questions • Which genes does DiplonemamtDNA contain? • How are genes encrypted? • How does Diplonema decrypt these genes? A B Question #1…
Mitochondrialgene complement • Conventionalgene set in Diplonema mtDNA: • 17 typical mt-genes • 15 protein-coding • 2 structural RNAs • no tRNA genes. Valach et al 2018, JBC 293 aChlamydomonasreinhardtii unconventional is mt-gene structure …
Fragmented mt genes in Diplonema • Genes in small pieces (‘modules’, length 60-350 nt) • up to 11 modules per gene • a single module per chromosome • modules are transcribed separately and then joined(trans-spliced). A B m3 mtDNA m2 m4 m1 A A m1 Module transcripts m3 m2 m4 AAAAAAAAAA m1 m2 m3 m4 mRNA Marande & Burger 2007, Science 318; Vlceket al 2010, Nucleic Acids Res 39 Dp. mt-genes are not the only cases…
Other cases of gene fragmentation 1Feagin 1992 NAR 20; Winkler 1979 J Bact;2Perez-Martinez 2001, J BiolChem; 3Chalker 2013, CSH PerspBiol; 4Chang/Landweber 2005, PNAS How are pieces put together in Dp?…
Module joining intermediates Multiple transcript intermediates observed in Diplonema mito (example: a three-module gene). m2 m3 m1 m1 m2 m3 m1 m3 m2 m1 m3 m2 m1 m1 m1 m1 m2 m2 m2 m2 m2 m2 m2 m2 m3 m3 m3 m3 m1 m1 m1 m1 m2 m2 m2 m2 m3 m3 m3 m3
Inferred joining process Parallelized end maturation and trans-splicing in Diplonema mito. m3 m1 m2 A A B Trans-cription m1 m2 m3 End processing m1 m2 m3 m1 m3 m2 m1 m2 m3 m1 m1 m1 m1 m2 m2 m2 m2 m2 m2 m2 m2 m3 m3 m3 m3 Module joining m1 m2 m3 Accuracy?…
Accurate mate selection in trans-splicing ...exonGT..intron..AGexon… Spliceosomal Module transcript are joined in correct order: Motifs typical for intron splicing absent; How do cognate modules ‘find’ each other? in silico & experimental analyses… m1 m2 m3 m1 m3 ‘Archaeal’ Group I m1 m2 m2 m3 m2 m3 Group II m1 m2 m3 21 /43
Modules have unusual terminal groups Transcript intermediates characterized via RNA circularization; observation: + RNA ligase + RNA ligase module PO4 module NO circularization OH module PO4 module circularization. + 5ʹ kinase / 3ʹ phosphatase + 5ʹ kinase Kiethega, Yan, Turcotte, Burger 2013, RNA Biol 10 Closer look at ends…
Modules’ terminal groups ctn’d Systematic characterization: Terminal groups define ‘ligatable’ module ends: PO4 PO4 PO4 PO4 5ʹ module 5ʹ module PO4 PO4 OH OH internal module internal module OH OH AAAAA-OH AAAAA-OH 3ʹmodule 3ʹmodule However, there are not just 3 modules of a single gene…
Trans-splicing - a jigsaw puzzle ~100 distinct module transcripts in Diplonemid mitochondria 17 accurately assembled transcripts; i.e. no swapping of modules within a given transcript; no mixing of modules from different genes; m1 m2 m3 m4 m1 m3 m2 m4 m1 m2 m3 m1 m2 m3 How ismatchmakingachieved? Wetestedseveralhypotheses…
Possible mate selection strategies Hypothesis 1: Complementarycis sequence motifs m1 m2 m2 m1 No support A dynamic programming algorithm finds the max. nr. of base pairs that can be formed between two modules at a same min. distance (O) from module ends, in real and simulated data No significant difference between real and simulated data. m1 m2 O L Kiethega, Yan, Turcotte, Burger 2013, RNA Biol10
Possible mate selection strategies ctn’d Hypothesis 2:module matchmaking guided by trans factors Trans factors could be guide RNAs or guide proteins. m1 m2 :::: m1 m2 :::::::::
Possible mate selection strategies ctn’d Hypothesis 2a:RNA trans factors guide module matchmaking Experimental test: RT-PCR, both topologies, for cox1 Low-abundance candidates found for only 5 out of 8 junctions; In silico test:Search with regular expressions segments in ‘omic sequence that can pair with two cognate modules, allowing a bridge between interacting elements. Topology can be collinear or permutated; Excessive numbers of predicted candidates found. m1 m2 5′ 3′ No support m1 m2 3′ 5′
A minimum of assumptions Hypothesis 3:trans-splicing is catalyzed by a complex (‘joinosome’) containing an RNA ligase. Candidate RNA ligases (nucleus-encoded) RNA ligases of ATP-grasp fold(type 1, type 2)2: 3ʹ-OH 5ʹ-PO4 RtcB-like RNA ligases: 3ʹ-OH 5ʹ-PO4 (corresponding to module ends) RtcB-ligase should be associated with guide; Isolate RtcB-containing mitochondrial complex. guide ligase OH PO4 m1 m2 1 Review: Burroughs & Aravind 2016, Nucl Acids Res ;2 Moreira et al 2015, BMC StructBiol 15
Joinosome isolation Work in progress: Express all three Diplonema RtcB genes in E. coli; ~ Raise antibodies against RNA ligases encoded in nuc. genome; immuno-precipitate joinosomes out of mitochondrial lysate; mass spectrometry, RNA-seq, DNA-seq of joinosomes. guide ligase OH PO4 m1 m2 Diplonema mt genes are not only fragmented…
Talk’s roadmap √1. What is known about mitochondria • genomes • gene expression • intron splicing 2. Unusual mitochondrial genome & expression in a micro-eukaryote: • diplonemids • broken genes & compensation • mutated genes & compensation 3. Evolutionary trends.
Mt genes sequences are mutated Compared to transcripts & proteins, Diplonema mt-genes have nucleotide deletions at module ends (≤ 30 nt) clustered nt substitutions inside modules (≤ 45/100 nt). transcript: m1/m2-junct: ..UCAGUCGUACUACAGCUCUUUUUUUUUACCAUAGCACCAACC... Gene rnlrnl-m1: ...TCAGTCGTACTACAGCTC rnl-m2:ACCATAGCACCAACC... transcript UGUGGUUUGUGUUGGAUGAUUUAUUGUAUUUGGUAGUGUAUUUGUUUUGGUUUU... gene nad4....CACAGCCTGTGCTGGATGACCCACCATACCTAACAGTACATCCACCTCGGCCTC... Moreira et al 2016, Nucleic Acids Res 44 Extent of mutations in mtDNA…
Mutations in Diplonemamt genes Closer look at types of mutations and theirparticular compensation…
Compensations by RNA editing Defects & compensations in Diplonema mt-genes: 1. Deletions at gene module ends Us appended at module transcript 3′-ends; Reminiscent of kinetoplastid RNA editing.. : module transcript : U-appendage sites nad3 nad9 nad2 nad6 nad4L
Kinetoplastid mt-gene expression Multiple chromosomes - Maxicircle: ‘cryptic’ protein genes (indels) - Minicircles: guide RNAs for RNA editing Leishmania Trypanosoma Crithidia Bodo insertion deletions Diplonemids Gene & primary transcript AGUUCAGGCCACGACCAGUGUCC guide RNA UCAGUCCGGUGCUGGUCACAGG AAAA AGUCAGGCGUUUCACGACCUAGUGUCC edited transcript UCAGUCCGUAAAGUGCUGGAUCACAGG endonuclease Editosome exonuclease TUTase others RNA ligase Review: Stuart et al 2005, Trends Biochem Sci39 ; Review: Gott & Emerson 2000, Annu Rev 34 Quitedifferentfrom U-based RNA editing in Diplonema mitos…
Mutation compensation by RNA editing Coding sequence defects in Diplonema mt-genes • 1. Deletions at gene module ends • Us appended at module transcript 3′-ends (distinct from kinetoplastid U-indels) • 2. Substitutions inside gene modules • deaminationsin module transcripts • C-to-U • A-to-I(Inosine; interpreted as G) • prior to module joining. : module : U-appendage sites : substitution sites nad3 nad9 nad2 nad6 nad4L
Experimental confirmation of Inosines Substrate: mt-SSU rRNA isolated from Diplonema ..UGUUAUUAGUUUGUGGUUCCAGUAU..pre-edited ..UGUUIUUIGUUUGUGGUUCCAGUAU..edited (G in RNA-Seq) 1. glyoxal/borate treatment (glyoxalatesGs but not Is or other nts) ..UGUUIUUIGUUUGUGGUUCCAGUAU.. 2. RNase T1 digestion (cuts RNA 3′ to Gs and Is, but not glyoxalated Gs) ..UGUUIUUIGUUUGUGGUUCCAGUAU.. 3. Primer extension (stops at first cleavage site) primerextension. RNA editing found in other organisms…
RNA editing across eukaryotes In nucleus (animals): • mostly A-to-I substitutions in mRNAs minor changes of translated protein; in organelles (outside animals): • C-to-U & U-to-C substitutions • any-to-any nucleotide (nt) substitutions at tRNAends • mono- and oligo-nt insertions& deletions (indels) • multiple U indels • critical for product’s function; unique to Diplonema: • A-to-I substitutions in mt-mRNAs, mt-rRNAs • U appendage. Acanthamoeba Dictyostelium Trypanosoma To summarize…
Summary - gene defects compensated • Defective genetic information in Diplonema mitochondria: • fragmented genes • deletions and substitutions in coding regions • Defects compensated post-transcriptionally Defects Gene fragmentation Nt deletions Nt substitutions Compensation trans-splicing of module transcript U-appendage RNA editing deamination RNA editing. Overly complicated system…
Rube-Goldberg gene expression? D E A c B A B D E C Source: Wikipedia Emergence?…
How it may have happened – molecular level Posited origin of multipartite mt-genome, fragmented genes, sequence mutations: 1. Transposable element invades mtDNA; 2. intramolecular recombination leads to sub-circles; 3. recombination events cause module loss and point mutations; 4. trans-splicing mends fragments; 5. U-appendage RNA-editing compensates for lost modules; 6. substitution RNA-editing compensates for point mutations. Invasion and propagation of mobile element Excision by recombination RNA trans-splicing & editing Evolutionary forces?…
How it may have happened – evolutionary mechanisms • Evolutionary sorting forces: • adaptive (natural selection) • fitness gain ?? damage control ?? • neutral (genetic drift) • neutral constructive evolution1 • pre-existing enzymes with trans-splicing and RNA editing activity… • …allowed mt-genome & gene fragmentation, and sequence mutations to happen; • accumulating defects make reversion unlikely. Invasion and propagation of mobile element Excision by recombination RNA trans-splicing & editing 1Corvello, Gray 1993, Trends Genet 9 ; Stoltzfuss 1999, MolEvol 49 ; Gray et al 2010, Science 330 Final outlook…
Conclusion & outlook • The unique machinery for Diplonema’s mitochondrial gene expression must have been recruited from preexisting components; • novelty emerged by evolutionary tinkering1; • Our goal: dissect novel molecular machineries to trace back components’ origin. 1François Jakob 1977 “Evolution and tinkering”, Science 196 The end.
Collaborators & funding Lab members involved in project Matus Valach (post-doc biochemistry) Sandrine Moreira (PhD program bioinformatics) Alexandra Léveillé-Kunst(MSc program biochemistry) Paul Stretenowich(MSc bioinformatics intern) LionnelLemogo (MSc bioinformatics intern) Wetlab: JA Gonzalez Alcazar, G Kiethega, S Breton, S Teijeiro, Y Yan, P Poliak, M Cool, M Smaali, K Yamini, W Marande. Drylab: A Breton, J Coquet, L Perras, S Prigent, T Mesquita, LA Le Minh, R Palvadeau. Collaborations Michael W. Gray (Dalhousie University, Halifax, Canada) B. Franz Lang (Université de Montréal, Canada) Julius Lukeš (University of South Bohemia, Czech Rep.) Marcel Turcotte (University of Ottawa, Canada)
Chromosome number and module distribution across diplonemids vary Mono-module Multi-module cox3-m1 chromosome Dp_A0x 1Kaur et al, in prep.
Mt-gene fragmentation conserved in D/R clade All but one gene breakpoint conserved in Diplonema & Rhynchopus clade D. a. D. p. D. sp. R. e. m1 m2 m1 m2 m1 m1 m2 : module
Diplonemids reign the oceans Among eukaryotes in sunlit global ocean, diplonemids rank 7 ! in abundance 3 ! sequence diversity (18SrRNA): ~12,300 ! Species. Nr. of Ref. Operational tax. Units based Barcodes on 18S rRNA sequence deVargas et al 2015, Science
Recurrent cis sequence motifs? Hypothesis 1a: Sequence motifs in modules or adjacent regions involved inmodule matchmaking. Test:look for recurrentsequencemotifsdefining ‘left’-side and ‘right’-side joining sites*. Approach: Search for consensus sequence motifsat all ‘left’ sites and all ‘right’ sites of module junctions from gene cox1 in four diplonemids. Tools: muscle1, weblogo2. m3 m3 m1 m2 *Catalytic RNA motifs (hammerhead, HDV, twister, twister sister) are characterized by both conserved sequence motifs and 2D structure. m1 m2 m2 Kiethega et al 2011, MolBiolEvol 28:2425; 1Edgar et l 2004, BMC Bioinformatics 5:113;2http:weblogo.Berkley.edu.lgo.cgi
No recurrent cis sequence motifs Hypothesis 1a: Sequence motifs in modules or adjacent regions involved inmodule matchmaking. Result: m3 m1 m2 m2 Kiethega et al 2011, MolBiolEvol 28:2425
Recurrent cis 2D motifs? Hypothesis 1b: 2D structure motifs in modules or adjacent regions involved in module matchmaking. Test:look for recurrentstructuremotifsdefining ‘left’-side and ‘right’-side joining sites*. Approach: Search for consensus 2D structure motifs shared by all ‘left’ ends and all ‘right’ ends, respectively, of module junctions from gene cox1. Tools RNAalifold1, RNAshapes2 etc. m1 m2 m3 m1 m2 m2 m3 *Catalytic RNA motifs (hammerhead, HDV, twister, twister sister) are characterized by both conserved sequence motifs and 2D structure. 1 Kiethega et al 2011, Mol Biol Evol 28:2425; 2ViennaRNA Package 2011 Algos4MolecBiol 6:26; 3Steffen et al 2006 Bioinformatics 22:500
No recurrent 2D cis motifs Hypothesis 1b: 2D structure motifs in modules or adjacent regions involved in module matchmaking. m1 m2 m2 m3