Skeletal Muscle I
This overview delves into the fundamental features of vertebrate skeletal muscle, highlighting its striated, polynuclear cells organized into sarcomeres. We explore the tissue structure, including how muscle fibers are surrounded by different connective sheaths and the role of specialized compartments like the sarcoplasmic reticulum. The mechanisms of contraction through sliding filaments are covered, alongside critical concepts in excitation-contraction coupling and neural control. Lastly, we discuss how motor neurons induce smooth, sustained muscle contractions through precise electrical signaling.


Skeletal Muscle I
E N D
Presentation Transcript
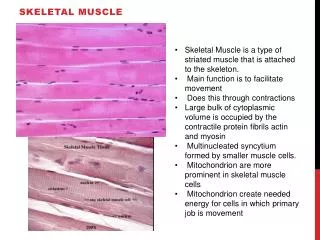
Basic Features of Vertebrate Skeletal Muscle • Filaments organized as sarcomeres (I.e. striated) • Cells large ( 200 micron diameter, up to meters long), polynuclear, formed by end-to-end fusion of myocytes during development. • Single, mononeuronal innervation (Ach, nicotinic receptors)
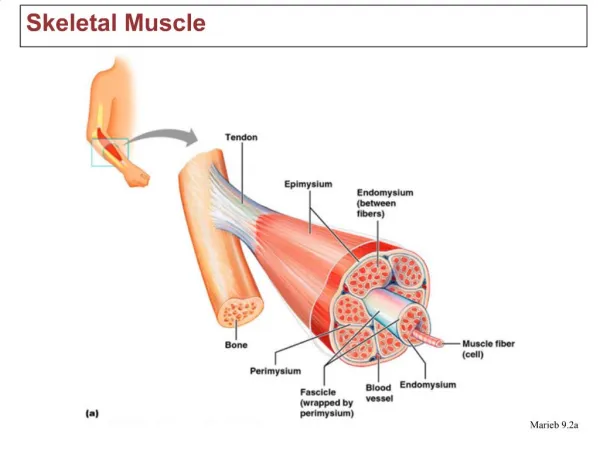
Tissue-level structure breakdown Muscle (consisting of many cells/fibers) bounded by epimysium Muscle fascicles (bundles) bound by perimysium Each fiber surrounded by endomysium = basal lamina which supports capillaries and axon terminals Each fiber has a sarcolemma=plasma membrane
Within the fiber there are two separate functional compartments: The cytoplasmic compartment containing several myofibrils composed of sarcomeres end-to-end, + mitochondria, etc. Surrounded by a network of hollow tubes of sarcoplasmic reticulum (a specialized endoplasmic reticulum)
This electron micrograph shows several complete sarcomeres of adjacent myofibrils
Major protein components of thin filaments • Actin: backbone + myosin binding sites • tropomyosin: regulates myosin access • troponin: binds Ca++ ; controls tropomyosin • nebulin: believed to serve as a scaffold for assembly of thin filaments and as a molecular yardstick that regulates their length.
The sarcomere is the basic functional unit of striated muscle This cartoon shows the thin-filament components listed in the previous slide and also shows titin, a protein that links the ends of thick filaments to the Z disc
Contraction of striated muscle involves sliding of thick and thin filaments. Implications of the sliding filament mechanism are seen in experiments in which the length-tension relationship of muscle is measured, as shown in the next slide.
The total tension is the sum of an active and a passive component. The active component is maximal when overlap of thick and thin filaments is optimal. Much of the elasticity of the passive muscle is now believed to be due to titin.
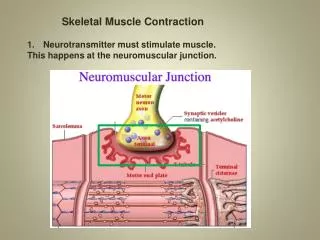
Points to remember • Acetylcholine is always the transmitter at the first synapse outside the CNS • Nicotinic receptors are always at the first synapse outside the CNS, are ionotropic and always excitatory. • In the parasympathetic pathway, acetylcholine is also the transmitter at the target organ, although other mediators may be coreleased (bombesin, for example). Parasympathetic target organs have muscarinic receptors for Ach (5 subtypes –all are metabotropic and may be excitatory or inhibitory) • The transmitter at sympathetic target organs is always norepinephrine; the receptors may be one or more of the four basic adrenergic receptor types (alpha1, alpha2, beta1, beta2 – these are all metabotropic and may be excitatory or inhibitory)
Skeletal Muscle Excitation: Facts • There is a 1:1 relationship between neuronal and muscle APs • Following an AP, a pulse of Ca++ in the muscle cytoplasm starts within a few msec and lasts for at least tens of msec - this causes the contractile machinery to become active. The [Ca++] achieved around the contractile machinery is enough to activate all (or almost all) of the contractile machinery. • The next slide shows the relationship between electrical excitation, Ca++ release, and tension development
In this experiment, the muscle is loaded with a probe whose fluorescence is quenched by Ca++. Notice that force development does not seem to begin until Ca++ levels have already begun to fall, and continues after [Ca++] has returned to baseline. This is the result of energy storage in the muscle’s series elastic elements.
Motor neurons talk to skeletal muscles in bursts of action potentials that result in smooth, sustained contractions • The next slide shows an experiment in which a twitch – the result of 1 AP – is compared to the tetanic contraction caused by a train of action potentials
The Ca++ pulse from 1 AP lasts longer than the electrical refractory period - allowing successive Aps to summate their contractile effects, giving a tetanus, or smooth contraction
The Problem Of Synchronizing the Contractile Machinery • the muscle AP spreads throughout the cell membrane within a few msec, BUT - how does an electrical signal at the cell surface cause myofibrils in the interior of the cell to become active? - fiber diameter is too great for a chemical signal crossing the cell membrane and diffusing to the central myofibrils to give the rapid onset and offset of contraction that we observe.
The Solutions • Transverse tubules of cell membrane penetrate the cell surface at intervals of 1 sarcomere length, carrying the electrical signal to the cell interior. • There is a store of Ca++ inside the SR. • T tubules pass close to the lateral cisternae of the SR; a T-tubular AP causes Ca++ release within the muscle fiber.
The square marks a muscle triad: a central T tubule with a lateral cisterna on each side.
The drawing shows an enlargement of the relationship between T tubule and lateral cisternae. Note the endfeet that seem to connect the two.
How does a T-tubular AP cause the SR to release Ca++? A pop-up hypothesis. Endfeet consist of 1. A voltage-sensitive channel protein in the T-tubular membrane (the dihydropyridine receptor), sitting above 2. a Ca++ channel in the SR membrane (the ryanodine receptor).
The DHP receptor serves as a voltage sensor that plugs the ryanodine receptor with its cytoplasmic domain when the T tubule is normally polarized, and lifts the stopper upon depolarization, allowing Ca++ to diffuse out down its concentration gradient.
The initial Ca++ release triggers additional Ca++ release Since only half of the endfeet have voltage sensors, it is currently believed that the initial Ca++ release triggers the opening of additional Ca++ release channels, a process called Ca++-induced Ca++ release.
The Ca++ switch for muscle contraction Tropomyosin- threads of tropomyosin lie along the thin filament. In the relaxed muscle each tropomyosin blocks a series of 8 binding sites for myosin heads. Troponin- a 3-subunit troponin controls each tropomyosin. As shown in the next slide, Ca++-binding to the C subunit of troponin causes a conformational change in the troponin that rolls or pulls tropomyosin away from the myosin binding sites.
Relaxation is the result of Ca++ recovery back into the SR by active transport - there is a Ca++-ATPase in the SR membrane. The level of activity of this enzyme determines twitch duration. Accumulation in the cisternae is aided by the presence of a Ca++-binding protein called calsequestrin that is present specifically within the cisternae.