Download

1 / 47
0 likes | 7 Views
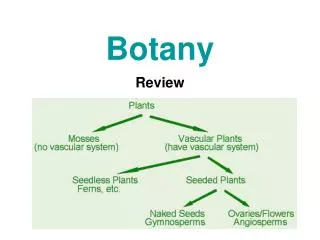
Dive into the essential concepts of Phanerogams and Paleobotany with this comprehensive PG TRB Botany Unit II study material from Professor Academy. Covering Pteridophytes, Gymnosperms, and Paleobotanical studies, the material includes detailed modules on classification, evolutionary theories, economic importance, fossil records, and stelar evolution. Ideal for postgraduate teaching aspirants preparing for TRB exams, this resource offers structured chapters, diagrams, memory tips, and model MCQs to ensure thorough preparation.<br><br>

E N D