Download

1 / 50
540 likes | 1.22k Views
2011 MECHANISMS OF CYTOPLASMIC mRNA TURNOVER IN EUKARYOTES. Kim Keeling, Ph.D. kkeeling@uab.edu Room 456, Bevill Bldg 975-6585. Lecture Overview. 1. General mRNA turnover pathways - Play key role in controlling basal gene expression levels 2. Aberrant RNA turnover pathways

E N D
2011 MECHANISMS OF CYTOPLASMIC mRNA TURNOVER IN EUKARYOTES Kim Keeling, Ph.D. kkeeling@uab.edu Room 456, Bevill Bldg 975-6585
Lecture Overview 1. General mRNA turnover pathways - Play key role in controlling basal gene expression levels 2. Aberrant RNA turnover pathways - Recognize and degrade aberrant mRNAs or mRNAs that are translated inefficiently - Increases the quality control of mRNA biogenesis and gene expression 3. Specialized mRNA turnover pathways - mRNA turnover acts as a site for regulation in response to specific signals (hormones, cell cycle, viral infection, differentiation, nutrient availability, stress) - AU-rich element mediated (ARE) 4. Specialized sites of cytoplasmic mRNA turnover - Processing bodies (P bodies) - Exosome granules for ARE-mediated mRNA turnover
Determining mRNA Stability Experimentally Northern blot of ACT1 mRNA 1. Shut off transcription: thiolutin, actinomycin D, promoters that can be repressed (such as copper or tetracycline regulated promoters). 2. Isolate mRNA at several time points after transcriptional shut-off. 3. Determine the percent of mRNA remaining at each time point after transcriptional shut-off. 4. Graph the level of RNA remaining for each time point on a semi-log graph. The slope = the rate of mRNA decay. The time at which half of the mRNA is degraded is the half-life of the mRNA. inhibit expression WT tpa1∆ time (m) 0 10 20 40 0 10 20 40 100 90 80 70 60 50 40 t1/2 = tpa1 24 min % ACT1 remaining 30 20 t1/2 = wild type 12 min 10 0 10 20 30 40 Time (m) Keeling et al. (2006) Mol Cell Biol 26:5237
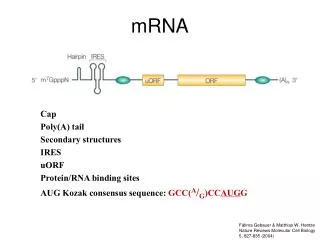
Nucleus Cytoplasm mRNAs Are Modified During Nuclear Processing to Promote Cytoplasmic Stability 1. Addition of 5’ cap and the CAP binding protein complex 2. Addition of the 3’ poly(A) tail and multiple poly(A) binding proteins 3. Circularization of mRNA by bridging the 5’ CAP and the 3’ poly(A) tail via eIF4G Goldstrohm & Wickens (2008) Nat Rev Mol Cell Biol 9:337
The Major Eukaryotic mRNA Turnover Pathways 5’ 3’ pathway 3’ 5’ pathway CAP hydrolysis Biochem. Soc. Trans. (2006) 34:35
The Major Eukaryotic mRNA Turnover Pathways 5’ 3’ pathway 3’ 5’ pathway CAP hydrolysis Biochem. Soc. Trans. (2006) 34:35
Mammalian Deadenylation Occurs in Two Phases PAN2 complex Slow CCR4 complex Fast Muhlemann (2005) Nat Struct Mol Biol 12:1024
Deadenylase Machinery 1) PAN complex: - First phase - Slow, distributive - Stimulated by PAB - Also trims poly(A) tails in nucleus 2) CCR4 complex: - Second phase - Fast, processive - Inhibited by PAB - Independent function as a transcription factor 3) PARN complex: - Used for specialized rapid deadenylation (ARE-mediated decay) in mammalian cells Parker & Song (2004) Nat Struct Mol Biol 11:121
T T T T T T T T T T Control RNA Transcriptional shut off Determining Deadenylation Rates Experimentally RNase H Northern blot RNase H + oligo dT poly(A)- PTETOFF AAAAAAAAAA Probe DNA oligo Northern blot slow synchronous fast asynchronous PAN complex CCR4 complex Yamashita et al. (2005) Nat Struct Mol Biol 12:1054
PAN and CCR4 Sequentially Deadenylate mRNAs Enhanced 2nd phase Enhanced 1st phase No difference Delayed 1st phase Delayed 2nd phase No difference Yamashita et al. (2005) Nat Struct Mol Biol 12:1054
Deadenylation Is Coupled to Translation Termination Hosoda et al (2003) JBC 278:38287 • - eRF3 is a GTPase that mediates translation termination. • - The N-terminal domain of eRF3 interacts with the Pab1p C-terminal domain. • - This interaction inhibits the ability of Pab1p molecules to oligomerize • efficiently on the poly(A) tail. This increases the exposure of the poly(A) • tail to deadenylases and turnover.
eRF3 Binds to Pab1p and Inhibits Its Oligomerization Hoshino et al. (1999) Biochem (Mosc) 64:1367 • - Gel shift assay: radio-labeled RNA is incubated with increasing • amounts of Pab1p protein. • - A shift in the mobility of the RNA indicates Pab1p protein binding. • - Addition of the eRF3 N-terminal domain to the binding assay • inhibits the ability of Pab1p to oligomerize on the RNA.
Hosoda et al (2003) JBC 278:38287 The eRF3-Mediated Inhibition of Pab1p Results in Deadenylaton and mRNA Turnover - Expression of a mutant eRF3 where the N-terminal domain is deleted so that it can no longer interact with Pab1p results in longer poly(A) tail length. This mutant also leads to an increased mRNA half-life = decreased turn- over rate. - This suggests that normally, eRF3 mediates mRNA turnover by decreasing Pab1p oligomerization on poly(A) tails which leads to deadenylation and mRNA decay. This represents one way to mediate general mRNA decay.
The Major Eukaryotic mRNA Turnover Pathways 5’ 3’ pathway Pan2, Ccr4 CAP hydrolysis Biochem. Soc. Trans. (2006) 34:35
Decapping Enzymes Dcp1 - Dcp2p = catalytic subunit - Dcp1p = stimulates Dcp2p - Hedls = also needed to stimulate Dcp2p activity in mammals - In yeast, Dcp1p and Dcp2p can directly interact with each other to stimulate decapping - Dcp2p is differentially expressed in mammalian tissues - Another decapping enzyme was identified in the cytoplasm of mammalian cells called Nudt16. No ortholog found in yeast, C. elegans or Drosophila. - Nudt16 and Dcp2 appear to each work on subsets of cellular transcripts. Hedls Yeast interactions Human interactions Dcp2 Simon et al. (2006) TIBS 31:241 Nudt16 Taylor & Peculis (2008) NARS 36:6021
Control of the Decapping Reaction • 1. Poly(A) tail status - Poly(A) binding protein inhibits decapping by physically interacting with the 5’ cap complex (via eIF4G) to stabilize the cap and enhance translation. 2. CAP status - The CAP protein complex must dissociate from the cap prior to decapping. 3. Translation status - A transition must take place from a translation competent state to a state where translation does not take place (change in mRNP structure). 4. Assembly of the decapping complex - Specific factors are required for the localization, assembly, and activation of the decapping complex.
Parker & Song (2004) Nat Struct Mol Biol 11:121 Decapping Activators Pat1p - Interacts with both poly(A) [+] and poly(A) [-] transcripts - Seeds decapping complex onto mRNA - Interacts with Dcp1p, Dcp2p, Lsm1-7p, Dhh1p, Xrn1p - Along with the Lsm complex, represses translation Lsm1-7p - 7 ring complex interacts with poly(A) [-] mRNA - Facilitates the assembly of the decapping complex in cytoplasmic and nuclear decapping - Interacts with Dcp1p, Dcp2p, Dhh1p, Pat1p, Xrn1p Dhh1p - ATP dependent helicase - Binds directly to mRNA substrate - Interacts with Dcp1p, Dcp2p, Lsm1-7p, Ccr4p, Pop2p, Edc1p, Edc2p Edc1/Edc2 - Binds directly to mRNA substrate - Enhances decapping activity - Interacts with Dcp1 and Dcp2 - Enhances alterations in mRNA stability in response to nutrient changes
The Major Eukaryotic mRNA Turnover Pathways 5’ 3’ pathway Pan2, Ccr4 Dcp2, Nudt16 CAP hydrolysis Biochem. Soc. Trans. (2006) 34:35
The Major Eukaryotic mRNA Turnover Pathways 5’ 3’ pathway Pan2, Ccr4 Dcp2, Nudt16 CAP hydrolysis Xrn2/Rat1 = nuclear exonuclease Biochem. Soc. Trans. (2006) 34:35
The Major Eukaryotic mRNA Turnover Pathways Pan2, Ccr4 CAP hydrolysis Biochem. Soc. Trans. (2006) 34:35
Exosome Components Core Subunits: all essential Rrp4p Rrp40p Csl4p Rrp41p/Ski6p Rrp42p Rrp43p Rrp44p/Dis3p Rrp45p Rrp46p Mtr3p Associated factors: Mtr4p essential ATP-dependent helicase Ski2p non-essential ATP-dependent helicase Ski3p non-essential TPR repeat Ski8p non-essential WD repeat Ski7p non-essential GTPase Nuclear subunits: Rrp6p non-essential nuclear subunit Parker & Song (2004) Nat Struct Mol Biol 11:121
The Exosome Gains Access to mRNAs for Degradation Via Ski7p and the SKI Complex - Ski7p bridges an interaction between the mRNA-bound SKI complex and the exosome. - In the nucleus, a group of factors called the TRAMP complex is the bridge between mRNAs and the exosome. Araki et al. (2001) EMBO J 20:4684
The Major Eukaryotic mRNA Turnover Pathways Pan2, Ccr4 Dcp2, Nudt16 CAP hydrolysis Biochem. Soc. Trans. (2006) 34:35
Final Step of mRNA Decay: Degrading the 5’ Cap - The scavenger decapping enzyme hydrolyzes di- and tri-phosphorylated CAPs to the mono form. - Substrates are less than 10 nucleotides in length. Parker & Song (2004) Nat Struct Mol Biol 11:121
Summary of the General Eukaryotic mRNA Decay Pathways Meyer et al. (2004) Crit Rev Biochem Mol Biol 39:197
Nuclear mRNA Decay Also Utilizes Components of the 5' 3' and 3' 5' mRNA Turnover Pathways Moore (2002) Cell 108:431 TRAMP Xrn2p Nuclear Yes Yes Xrn2p/Rat1p TRAMP
2) Aberrant mRNA Decay Pathways A. Nonsense-mediated mRNA decay (NMD) - Degrades mRNAs with premature stop codons B. Nonstop mRNA decay (NSD) - Degrades mRNAs without a stop codon C. No-go mRNA decay (NGD) - Degrades mRNAs that have a stalled ribosome D. Ribosome extension-mediated decay (REMD) - Degrades mRNAs where ribosome translates past the stop codon and into the 3’ UTR
Nonsense-Mediated mRNA Decay - Specialized pathway that degrades mRNAs thatcontain premature translation termination signals - Protects the cell from translating mRNAs that might produce truncated peptides that could lead to harmful dominant negative effects - Occurs in all eukaryotes. - 30% of disease-generating mutations result in premature stop codons - Up to 10-20% of the transcriptome is regulated by NMD - PTC-containing transcripts caused by point mutations, frameshift mutations, mRNAs with faulty alternative splicing, pre-mRNAs that escape nuclear retention, mRNAs that contain upstream open reading frames, mRNAs that carry introns in 3´ untranslated regions, or mRNAs with long 3´ untranslated regions Czapllinski et al. (1999) Bioeassay 21:685
Splicing-dependent NMD Normal Termination Aberrant Termination Current NMD Models Splicing-independent NMD Wen & Brogna (2008) Biochem Soc Trans 36:514
core NMD components NMD Factors Associate With the EJC Core NMD Components: UPF3: associates with the EJC in the nucleus UPF2: perinuclear and binds to Upf3 as the mRNA is exported UPF1: associates at the stop codons in mRNAs during translation
Regulation of NMD by Upf1p Phosphorylation 1. The SURF complex composed of SMG1, UPF1, and the eRF1 and eRF3 release factors forms. This complex recognizes stop codons. 2. If the SURF complex interacts with a downstream EJC complex that includes UPF2, UPF3, and Y14 (the DECID complex), then SMG-1 phosphorylates UPF1. This marks the stop codon as premature. 3. UPF1 phosphorylation induces a change in the mRNP structure that recruits SMG5, SMG6, SMG7, and the PP2A phosphatase to dephosphorylate UPF1. This signals for the mRNA to be degraded. Kashima et al. (2006) Gene Dev 20:355
Aberrant Translation Termination Mediates NMD Muhlemann (2008) Biochem Soc Trans 36:497
Nonstop mRNA Decay - Pathway to degrade mRNAs thatdo not contain any stop codons - Typically caused by the presence of cryptic poly(A) addition sites that leads to polyadenylation of transcripts upstream of the termination signal - Utilizes Ski7p, the SKI complex, and the exosome to degrade nonstop messages Maquat (2002) Science 295:2221
No Go mRNA Decay - Degrades messages with translation elongation stalls (i.e. hairpin structures, pseudoknots, rare codons) - Requires two factors that bind to the A site of the stalled ribosome -- Dom34p (eRF1-like homologue) -- Hbs1p (eRF3-like homologue) - Promotes ribosomal subunit dissociation and peptidyl tRNAs to remove them from translation elongation stalls and recycle them - mRNA cleaved at stalled position by Dom34p which has endonuclease activity and then degraded by the exosome and/or Xrn1p Tollervey (2006) Nature 440:425
Pathway A site binder GTPase NMD eRF1 eRF3 NGD DOM34 HSB1 NSD ? SKI7 Many of the Specialized mRNA Decay Pathways Recruit Similar Proteins to the Ribosomal A Site to Trigger different mRNA Decay Mechanisms. Clement & Lykke-Anderson (2006) Nat Struct Mol Biol 13:299 Chen et al. (2010) Nat Struct Mol Biol 17:1233
Ribosome Extension-Mediated mRNA Decay - Constant Spring (CS) mutation in -globin changes the UAA stop codon at the end of the mRNA into a CAA (glutamine) codon - Mutation is the most prevalent non-deletion mutation that causes -thalassemia - Ribosome continues to translate 31 codons in the 3’ UTR until a stop codon is encountered (UAA) - Level of mRNA is severely decreased due to this mutation - CS mutation causes a decrease in the mRNA half-life due to rapid induction of deadenylation - Mechanism behind this rapid deadenylation is still unknown - REMD is cell-type restricted Kong & Liebhaber (2007) NSMB 14:670
Summary of mRNA Surveillance Pathways Doma & Parker (2007) Cell 131:660
3) Specialized mRNA turnover pathways - ARE-mediated mRNA decay
ARE-Mediated mRNA Turnover Goldstrohm & Wickens (2008) Nat Rev Mol CellBiol 9:337 ARE - AU rich elements (50-150nts) AUUUA; UUAUUUA(U/A)(U/A); or U-rich - cis-acting element located in 3' UTRs of mRNAs - transcripts that encode proteins that require rapid changes in response to stimuli such changes in the cell cycle, growth factors, response to microorganisms, inflammatory stimuli, and environmental factors - 10% of mammalian mRNAs contain AREs - A diverse set of trans-acting proteins bind to AREs. These proteins can mediate other protein interactions that modulate mRNA stability. Various ARE-associated proteins can promote rapid mRNA turnover by promoting enhanced decapping, deadenylation, exosome recruitment, endonucleolytic cleavage or combinations of these. Alternatively, some proteins that bind to AREs can stabilize the mRNA.
Various Triggers Alter the Balance of Effectors to Modulate ARE-mediated mRNA Decay Eberhardt et al. (2007) Pharm Ther 114:56
Example of ARE-Mediated Change in mRNA Levels Before and After the DNA Damage Response Under non-damage conditions: - AUF1 competes with the PABP for poly(A) tail binding, exposing it to PARN; TTP (tristetraproline) and KSRP (KH-type splicing regulatory protein) recruit PARN and CCR4 to deadenylate prior to degradation by the exosome. Under DNA damage conditions: - Genes involved in the DNA damage response pathway are up-regulated. HuR is up-regulated and competes with AUF1 for binding to the same ARE region. Loss of AUF1 binding stabilizes PABP association with the poly(A) tail. HuR also competes with TTP and KRSP to prevent recruitment of the deadenylases and exosome. Cevhar & Kleiman (2010) WIRE 1:193
4) Specialized sites of cytoplasmic mRNA turnover - Processing bodies - Exosome granules
RNA Processing Bodies (P bodies) • - Discrete cytoplasmic granular structures that contain a reservoir of 5’ 3’ • mRNA decay factors; NMD factors; RNA-induced silencing complex • - Found in all eukaryotes • - Size and number of P bodies depend upon the amount of RNA to be degraded • Conditions that promote P body formation include: • - Glucose deprivation • - Osmotic stress • - UV light • - Decreased translation initiation rates • - Non-translating mRNA • Note that all of these conditions involve moving mRNAs from a translatable pool that is ribosome-associated to a non-translatable pool that is not ribosome- associated • Conditions that reduce P bodies include: • - Inhibition of translation elongation (ribosome can’t dissociate from mRNA) • - RNase A treatment (RNA degraded) • - Increased rates of translation initiation (increase in ribosome-bound mRNA)
Identification of Factors That Reside in P Bodies Factors Not Found in P Bodies - Translation initiation factors - Ribosomal subunits - SKI proteins & exosome Kulkarni et al. (2010) Biochem Soc Trans 38: 242
RNA Decay Intermediates Localize to P Bodies - PolyG tract (18) in 3’ UTR blocks Xrn1p to create a mRNA decay intermediate - MS2 = bacteriophage coat protein binding site - MS2-GFP protein will bind to MS2 sites in mRNA and allows its localization by visualization of the green fluorescence protein (GFP) Sheth & Parker (2003) Science 300:805
P Bodies Disassemble After Translation Restoration - Polysome profile (ribosomes fractionated through a sucrose gradient) - Active translation = (+) polysomes - Inactive translation = (-) polysomes - P body-associated mRNAs can return to the translatable pool Brengues et al. (2005) Science 310:486.
Exosome Granules Are Distinct From P Bodies and Contain ARE mRNAs and PARN exosome protein merge P body protein exosome protein PARN merge ARE mRNA Lin (2007) JBC 282:19958
Lecture Overview 1. General mRNA turnover pathways - 5’3 & 3’5’ - Deadenylases - Decapping complex - Xrn1, exosome, DcpS 2. Aberrant RNA turnover pathways - Premature stop codons: nonsense-mediated mRNA decay (NMD) - No stop codons: non-stop mRNA decay (NSD) - Elongation stall: no-go mRNA decay (NGD) - Translation into the 3’ UTR: ribosome extension-mediated mRNA decay (REMD 3. Specialized mRNA turnover pathways - ARE-mediated mRNA turnover: AU-rich elements in the 3’ UTR are bound by proteins that modulate the stability of mRNAs in response to regulatory signals 4. Locale of mRNA turnover - P bodies: contain 5’3’ mRNA turnover machinery and degrade mRNAs that are no longer available for translation - Exosome granules: contain PARN and exosome and participate in ARE- mediated decay