Download

1 / 21

E N D
booksmedicos.org 7 C A P Í T U L O Glándula tiroides David S. Cooper, MD y Paul W. Ladenson, MA (Oxon.), MD La glándula tiroides es el único órgano de mayor tamaño del cuerpo, especializado para la producción de hormona endocrina. Su función es secretar una cantidad apropiada de las hormonas tiroideas, princi- palmente 3,5,3′,5′-l-tetrayodotironina (tiroxina, T4), y una menor cantidad de 3,5,3′-l-triyodotironina (T3), que surge principalmente a partir de la desyodación extratiroidea subsiguiente de T4. En los teji- dos blanco, la T3interactúa con receptores de T3 nucleares que, a su vez, están unidos a secuencias de nucleótidos especiales en las regiones promotoras de los genes que están regulados de manera positiva o negativa por la hormona tiroidea. Entre sus acciones para sostener la vida, las hormonas tiroideas promueven el crecimiento normal del feto y durante la niñez y el desarrollo del sistema nervioso central; regulan la frecuencia cardiaca y la contracción y relajación miocárdi- cas; afectan la motilidad gastrointestinal y la depuración renal de agua, y regulan el gasto de energía, la generación de calor, el peso y el metabolismo de los lípidos del cuerpo. Además, la tiroides contiene células parafoliculares o C que producen calcitonina, un polipéptido de 32 aminoácidos que inhibe la resorción ósea, pero que no tiene un papel fisiológico manifiesto en seres humanos. Sin embargo, la calci- tonina es importante en clínica como un marcador tumoral produci- do por cánceres medulares de la tiroides que surgen a partir de estas células (capítulo 8). palpable en estados o enfermedades que causan inflamación difusa de la tiroides o estimulación de esta última (figura 7-1). El crecimiento hacia arriba de la glándula tiroides está limitado por la fijación del músculo esternohioideo al cartílago tiroides. Sin embargo, no hay impedimento para el crecimiento en dirección pos- terior y hacia abajo, de modo que el agrandamiento de la tiroides, o bocio, a menudo se extiende en dirección posterior e inferior, incluso hacia la parte superior del mediastino (bocio retroesternal). La glándula tiroides tiene relaciones anatómicas importantes en clínica con los nervios laríngeos recurrentes, que tienen una trayecto- ria por detrás de la glándula, y dos pares de glándulas paratiroides que por lo general yacen detrás de las porciones superior y media de los lóbulos tiroideos. La tiroides también envuelve la tráquea y los már- genes posteriores de sus lóbulos colindan con el esófago; todas estas estructuras pueden quedar comprimidas por agrandamiento de la glándula, invadidas por enfermedades malignas de la tiroides, o lesio- nadas en el transcurso de una intervención quirúrgica tiroidea (figura 7-2). Dado que la cápsula tiroidea posterior está unida a la fascia pretraqueal, la glándula normalmente asciende y desciende con la deglución, lo que facilita su inspección y palpación. La tiroides tiene un rico aporte sanguíneo (figura 7-3), que puede estar aumentado en el hipertiroidismo, lo que da lugar a un ruido silbante audible (soplo) o incluso una vibración palpable (frémito). Desde el punto de vista microscópico, los tirocitos forman esferas huecas (folículos) con una luz central que contiene una agregación de tiroglobulina yodada denominada coloide y que representa las reser- vas hormonales de la glándula (figura 7-4). EMBRIOGENIA, ANATOMÍA E HISTOLOGÍA La glándula tiroides se origina en el embrión como una invaginación mesodérmica en el piso faríngeo formándose un agujero ciego, a partir del cual desciende en posición anterior a la tráquea y se bifurca, lo que forma dos lóbulos laterales, cada uno de los cuales mide aproximadamente 4 cm de longitud, 2 cm de ancho, y 1 cm de grosor durante la adultez. Puede haber tejido tiroideo ectópico en cualquier lugar a lo largo de este conducto tirogloso o más allá, desde la base de la lengua (tiroides lingual) hasta el mediastino. El conducto tirogloso también puede dar lugar a quistes en la línea media revestidos con epitelio escamoso, que pueden permanecer asintomáticos, o dar lugar a infecciones o tumores tiroideos. El extremo caudal del conducto tirogloso forma el lóbulo piramidal de la tiroides, que puede hacerse FISIOLOGÍA ESTRUCTURA Y SÍNTESIS DE HORMONAS TIROIDEAS Las hormonas tiroideas son tironinas yodadas, que comprenden dos porciones de tirosina unidas por un enlace éster (figura 7-5). Las células foliculares de la glándula tiroides están capacitadas para sinte- tizar la proteína precursora hormonal tiroglobulina, concentrar yo- duro dentro de la célula desde la circulación, y expresar un receptor 163 http://booksmedicos.org 07Gardner(163-226).indd 163 07Gardner(163-226).indd 163 26/8/11 17:22:27 26/8/11 17:22:27
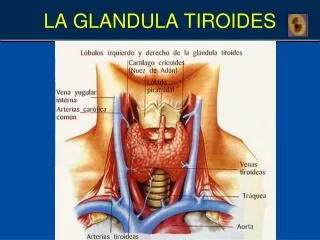
booksmedicos.org 164 CAPÍTULO 7 Glándula tiroides Hueso hioides Músculo omohioideo Cartílago tiroides Músculo esternohioideo Lóbulo piramidal Músculo cricotiroideo Glándula tiroides Músculo esternocleidomastoideo Tráquea Músculo esternotiroideo FIGURA 7–1 Anatomía macroscópica de la glándula tiroides del ser humano (vista anterior). que se une a la hormona estimulante de la tiroides (tirotropina, TSH), que promueve el crecimiento y las funciones biosintéticas de los tirocitos. bacterianos yodóforo usados en la recolección de la leche, colorantes de alimentos y mariscos. Los tirocitos expresan de manera abundante el cotransportador unidireccional de sodio-yoduro (simportador de Na-I–; NIS), que abarca las membranas basales de las células y transporta de manera activa yoduro desde la sangre. La glándula tiroides concentra y usa para la síntesis de hormona sólo una fracción del yoduro que se le proporciona, y el resto vuelve al fondo común en el líquido extrace- lular. En consecuencia, la captación fraccional normal de yoduro, que puede cuantificarse con un rastreador de yodo radiactivo, es de aproximadamente 10 a 30% después de 24 h. Debido a este mecanis- mo de concentración activo y la organificación subsiguiente del yo- duro intracelular, el fondo común de yodo intratiroideo es muy grande, de 8 a 10 mg en forma de hormonas tiroideas y tirosinas yodadas almacenadas. Este yodo proporciona un amortiguador en caso de deficiencia temporal de yodo en la dieta. METABOLISMO DEL YODO El yodo1 es un componente estructural clave de las hormonas tiroi- deas. En consecuencia, es un micronutriente esencial que se consume en los alimentos o el agua como yoduro o yodato, que se convierte en yoduro en el estómago. La Organización Mundial de la Salud (OMS) recomienda una ingestión diaria de yodo en la dieta de 150 μg para adultos, 200 μg para embarazadas y mujeres que amamantan, y 50 a 250 μg para niños. Dado que casi todo el yoduro se excreta por los riñones, la excreción urinaria de yoduro es un excelente índice de ingestión en la dieta. Durante milenios, el yodo se ha disuelto del suelo en muchas regiones montañosas y tierra adentro del mundo. En consecuencia, de acuerdo con la OMS, la deficiencia de yodo en la dieta, definida como una ingestión diaria de menos de 100 μg/día afecta a un esti- mado de 2 000 millones de personas, que es alrededor de una tercera parte de la población del mundo. Cuando la ingestión de yoduro es de menos de 50 μg/día, una tiroides de tamaño normal no puede sostener una producción suficiente de hormona, con agrandamiento resultante de la glándula (bocio) y, finalmente, hipotiroidismo. Las consecuencias de la deficiencia endémica de yodo en la dieta son en especial devastadoras para el feto y los niños en desarrollo, que requieren hormona tiroidea para el desarrollo neurológico y creci- miento normales. En la dieta norteamericana, el yodo se deriva principalmente de la sal yodada, conservadores yodato en alimentos horneados, productos lácteos que contienen rastros de agentes anti- SÍNTESIS Y SECRECIÓN DE HORMONA TIROIDEA La síntesis de T4 y T3 por la glándula tiroides comprende seis pasos principales: 1) transporte activo de yodo a través de la membrana basal hacia la célula tiroidea (atrapamiento); 2) oxidación de yoduro y yodación de residuos tirosilo en la tiroglobulina (organificación); 3) enlace de pares de moléculas de yodotirosina dentro de la tiroglobu- lina para formar las yodotironinas T3 y T4 (acoplamiento); 4) pinoci- tosis y después proteólisis de la tiroglobulina con liberación de yodo- tironinas y yodotirosinas libres hacia la circulación; 5) desyodación de yodotirosinas dentro de la célula tiroidea, con conservación del yodu- ro liberado para volver a usarlo, y 6) 5′-desyodación intratiroidea de T4 hacia T3. La síntesis de hormona tiroidea requiere que el NIS, la tiroglobu- lina y la enzima peroxidasa tiroidea (TPO) estén presentes, sean 1 En este capítulo, las palabras “yodo” que se refiere al elemento sin carga, y “yoduro”, que se refiere a su forma anión con carga negativa, se usan como sinónimos. http://booksmedicos.org 07Gardner(163-226).indd 164 07Gardner(163-226).indd 164 26/8/11 17:22:28 26/8/11 17:22:28
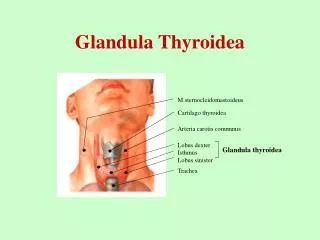
booksmedicos.org CAPÍTULO 7 Glándula tiroides 165 Istmo de la glándula tiroides Tráquea Esófago Lóbulo izquierdo de la glándula tiroides Nervio laríngeo recurrente Lóbulo derecho de la glándula tiroides Arteria carótida primitiva Nervio vago Músculo largo del cuello Vena yugular interna Fascia prevertebral Glándula paratiroides FIGURA 7–2 Corte transversal del cuello a nivel de T1, que muestra las relaciones de la tiroides. (Reproducida, con autorización, de Lindner HH. Clinical Anatomy. McGraw-Hill; 1989.) funcionales y no estén inhibidos. Este proceso se resume en las figuras 7-6 y 7-7. sólo cuatro sitios tirosilo se encuentran orientados de manera estérica para la hormonogénesis efectiva. El contenido de yodo de la tiroglo- bulina puede variar desde 0.1 hasta 1% por peso. Por ejemplo, en la tiroglobulina que contiene 0.5% de yodo, hay aproximadamente tres moléculas de T4 y una molécula de T3. El gen tiroglobulina, que reside en el brazo largo del cromosoma 8, contiene aproximadamente 8 500 nucleótidos, que codifican para el monómero proteínico pretiroglobulina, incluso un péptido señal de 19 aminoácidos. La TSH regula la expresión del gen tiroglobulina. Tiroglobulina La tiroglobulina es una molécula glucoproteica grande (peso mo- lecular [MW] de 660 000 kDa), compuesta de dos subunidades, cada una de las cuales contiene 5 496 aminoácidos. La tiroglobulina inclu- ye aproximadamente 140 residuos tirosilo, pero en cada molécula Vena yugular interna Vena tiroidea superior Arteria tiroidea superior Vena tiroidea media Arteria tiroidea inferior Arteria carótida primitiva Arteria y vena subclavias Nervio vago Vena tiroidea inferior Arteria tiroidea media (arteria de Neubauer) FIGURA 7–3 Arterias y venas relacionadas con la glándula tiroides. (Reproducida, con autorización, de Lindner HH. Clinical Anatomy. McGraw-Hill; 1989.) http://booksmedicos.org 07Gardner(163-226).indd 165 07Gardner(163-226).indd 165 26/8/11 17:22:28 26/8/11 17:22:28
booksmedicos.org 166 CAPÍTULO 7 Glándula tiroides FIGURA 7–4A. Tiroides normal de rata. Una capa única de células epiteliales cúbicas rodea el material PAS-positivo en el espacio folicular (coloide). Las células de mayor tamaño que se tiñen de color más claro, indicadas por las flechas, (I) son células C que producen calcitonina. Las células foliculares forman una capa epitelial que rodea el coloide (F). B. Tiroides inactiva de rata, varias semanas después de hipofisectomía. La luz de los folículos es de mayor tamaño, y las células foliculares más planas. C. Tiroides de rata, bajo estimulación intensiva con TSH. El animal fue alimentado con una dieta con deficiencia de yodo, y se le inyectó propiltiouracilo durante varias semanas. Hay poco coloide visible. Las células foliculares son altas y cilíndricas. Varias mitosis (m) son visibles. Reproducida, con autorización, de Halmi NS. En: Greep RO, Weiss L, eds. Histology. McGraw-Hill; 1973.) Después de que el mRNA que codifica para tiroglobulina se traduce en el retículo endoplasmático rugoso (RER), la proteína es glucosila- da durante el transporte por el aparato de Golgi (figura 7-7), donde dímeros de tiroglobulina se incorporan en vesículas exocíticas. A continuación, estas vesículas se fusionan con la membrana basal api- cal de la célula, desde la cual se liberan hacia la luz folicular. Ahí, en el borde apical-coloide, los residuos tirosina en la tiroglobulina son yodados. I HO CH2CH COOH NH2 3-Monoyodotirosina (MIT) I HO CH2CH COOH NH2 I 3,5-Diyodotirosina (DIT) CIO4−, SCN− bloquean el transporte activo de I− Los fármacos antitiroideos bloquean la yodación de la tiroglobulina I I HO O CH2CH COOH NH2 I I Na+ I− Na+ K+ Na+ 3,5,3′,5′-Tetrayodotironina (tiroxina [T4]) I Organificación: síntesis de T3 + T4 y almacenamiento en la tiroglobulina I− Na+ K+ I− I TPO HO O CH2CH COOH Coloide Suero Célula tiroidea NH2 I 3,5,3′-Triyodotironina (T3) I I Membrana basal (simportador Na+/I−) Microvellosidades apicales (pendrina I−, transportador de Cl−) HO O CH2CH COOH FIGURA 7–6 El transportador de yoduro en la célula tiroidea. El círculo oscuro grande representa el simportador de Na+/I– que transporta de manera activa I– hacia la célula; el círculo de color azul grande representa la Na+-K+ ATPasa que proporciona el gradiente de iones que impulsa la reacción. El I– es transportado a través de la membrana apical por la pendrina. La síntesis de hormona tiene lugar en el coloide-membrana apical, catalizada por la tiroperoxidasa (TPO). NH2 I 3,3′,5′-Triyodotironina (T3 reversa [rT3]) FIGURA 7–5 Estructura de hormonas tiroideas y compuestos rela- cionados. (Reproducida, con autorización, de Murray RK, et al. Harper’s Biochemistry. 24th ed. McGraw-Hill; 1996.) http://booksmedicos.org 07Gardner(163-226).indd 166 07Gardner(163-226).indd 166 26/8/11 17:22:28 26/8/11 17:22:28
booksmedicos.org CAPÍTULO 7 Glándula tiroides 167 Coloide Yodación de tiroglobulina Resorción de tiroglobulina Lisosomas Digestión por enzimas lisosómicas Oxidación de yoduro Incorporación de galactosa T3, T4 Incorporación de manosa Síntesis de proteína Síntesis de enzimas lisosómicas Transportador de yoduro Aminoácidos Yoduro T3 T4 Luz capilar FIGURA 7–7 Procesos de síntesis y yodación de la tiroglobulina (izquierda) y su resorción y digestión (derecha). Estos eventos ocurren en la misma célula. (Reproducida, con autorización, de Junqueira LC, Carneiro J, Kelley R. Basic Histology. 7th ed. McGraw-Hill; 1992.) Transporte de yoduro El yoduro (I–) es transportado a través de la membrana basal de los tirocitos por el NIS. El NIS unido a membrana, que obtiene su ener- gía a partir de una Na+-K+-ATPasa, permite que la glándula tiroides de ser humano mantenga una concentración de yoduro libre 30 a 40 veces más alta que la que hay en el plasma. La acción del NIS es esti- mulada desde el punto de vista fisiológico por la TSH, y fisiopatoló- gico por el anticuerpo estimulador del receptor de TSH propio de la enfermedad de Graves. Aunque los tejidos salival, gástrico y mamario expresan NIS y concentran yodo en menor grado que la tiroides, estos otros tejidos no organifican yoduro ni lo almacenan, y la TSH no estimula sus actividades de NIS. Grandes cantidades de yoduro supri- men tanto la actividad de NIS como la expresión del gen NIS, lo que representa mecanismos de autorregulación del yodo (véase más ade- lante). El ion perclorato (ClO4 el perclorato se ha usado para tratar hipertiroidismo, y tiene el poten- cial de ser un inhibidor ambiental de la función tiroidea. El NIS también puede concentrar pertecnetato (TcO4 roideas, lo que facilita el uso del radionúclido pertecnetato de sodio (Tc99mO4 ción de su actividad de atrapamiento. En el borde apical del tirocito, una segunda proteína de transpor- te de yoduro, la pendrina, transporta yoduro hacia la interface membrana-coloide, donde se convierte en un sustrato para la hormo- nogénesis tiroidea (figuras 7-6 y 7-7). Las mutaciones en el gen pen- drina (PDS o SLC26A4) que alteran su función causan un síndrome de bocio y pérdida de la audición adquirida durante la lactancia o en etapas tempranas de la niñez (síndrome de Pendred). Peroxidasa tiroidea La peroxidasa tiroidea (TPO), una glucoproteína unida a membrana (MW de 102 kDa) que contiene una porción hem, cataliza tanto la oxidación de yoduro como el enlace covalente de yodo a los residuos de tirosina de la tiroglobulina. La TSH estimula la expresión del gen TPO. Después de que se sintetiza TPO en el RER, se inserta en las membranas de las cisternas del RER, desde las cuales se transfiere hacia la superficie celular apical mediante elementos de Golgi y ve- sículas exocíticas. Ahí, en la interface célula-coloide, la TPO facilita tanto la yodación como el acoplamiento de los residuos de tirosina dentro de la tiroglobulina. –) compite con el yoduro por el NIS; Yodación de la tiroglobulina Dentro de la célula tiroidea, en la interface apical-coloide, el yoduro es oxidado con rapidez por peróxido de hidrógeno producido local- mente en una reacción catalizada por TPO; el intermediario yoduro activo resultante se une a residuos de tirosilo en la tiroglobulina. El peróxido de hidrógeno requerido probablemente es generado por una NADPH oxidasa en presencia de cationes de calcio, un proceso tam- bién estimulado por la TSH. La TPO también puede catalizar la yo- dación de moléculas de tirosilo en proteínas que no son la tiroglobu- –) hacia las células ti- –) para la visualización de la glándula tiroides y cuantifica- http://booksmedicos.org 07Gardner(163-226).indd 167 07Gardner(163-226).indd 167 26/8/11 17:22:29 26/8/11 17:22:29
booksmedicos.org 168 CAPÍTULO 7 Glándula tiroides lina, como la albúmina o fragmentos de tiroglobulina, pero no surgen hormonas activas a partir de estas proteínas. NH CO COOH CH I CH2 I HO Acoplamiento de residuos de yodotirosilo en la tiroglobulina La TPO también cataliza el acoplamiento de residuos de yodotirosilo en la tiroglobulina. Se cree que este es un proceso intramolecular que comprende oxidación de dos residuos de yodotirosilo acercados uno a otro por las estructuras terciaria y cuaternaria de la tiroglobulina, su enlace como un intermediario quinol éster, y división del quinol éster para formar una yodotironina (figura 7-8). Dentro de la molécula de tiroglobulina, dos moléculas de diyodotirosina (DIT) se acoplan para formar T4, y moléculas de monoyodotirosina (MIT) y DIT se aco- plan para formar T3. Los fármacos tiocarbamida, entre ellos metimazol, carbimazol y propiltiouracilo (PTU), son inhibidores competitivos de la TPO. Su capacidad resultante para bloquear la síntesis de hormona tiroidea (figura 7-9) los hace útiles en el tratamiento del hipertiroi- dismo. Tg HO I CH2 CH NH CO I NH2 1. TPO + H2O2 NH CO COOH CH I CH2 I O Tg O I CH2 CH NH CO I NH2 2. NO ENZIMÁTICA Proteólisis de tiroglobulina y secreción de hormona tiroidea En la figura 7-7 se ilustran los procesos de proteólisis y secreción de tiroglobulina en la hormonogénesis tiroidea. En la membrana apical del tirocito, el coloide queda rodeado en vesículas mediante pinoci- tosis, y es absorbido hacia la célula. A continuación, lisosomas que contienen enzimas proteolíticas se fusionan con la vesícula de coloi- de, esto libera T4 y T3, así como yodotirosinas inactivas, péptidos y aminoácidos individuales. Las hormonas tiroideas que tienen activi- dad biológica, T4 y T3, entran a la circulación; la DIT y la MIT son desyodadas, y su yoduro se conserva. La secreción de hormona tiroi- dea es estimulada por la TSH e inhibida por el exceso de yoduro (véase más adelante) y litio. En circunstancias normales, la célula tiroidea libera una pequeña cantidad de tiroglobulina intacta, y circula en la sangre. La concentración sérica de tiroglobulina está notoriamente aumentada en varias enfermedades de la tiroides, entre ellas tiroiditis, bocio nodular y enfermedad de Graves. Dado que casi todas las enfermedades malignas que surgen a partir del epitelio tiroideo, como los cánceres papilar y folicular de la tiroides tam- bién sintetizan tiroglobulina, ésta es un útil marcador tumoral circu- lante. NH CO COOH C H I CH2 Intermediario quinol éter I O Tg O I CH2 CH NH CO I NH2 3. REORDENAMIENTO NH CO COOH C Dehidroalanina I CH2 I HO Tg O I CH2 CH NH CO T4 I NH2 FIGURA 7–8 Esquema de acoplamiento hipotético para la forma- ción intramolecular de T4 dentro de la molécula de tiroglobulina. Está indicado el principal sitio hormonogénico en el residuo de tirosilo 5. (Reproducida, con autorización, de Taurog A. Thyroid hormone synthe- sis. Thyroid iodine metabolism. En Braverman LE, Utiger RD, eds. Werner and Ingbar’s The Thyroid. 7th ed. Lippincott; 1996.) Desyodación intratiroidea La MIT y la DIT que se forman durante la síntesis de hormona tiroi- dea son desyodadas por la desyodasa intratiroidea, una flavoproteína dependiente de NADPH que se encuentra en las mitocondrias y los microsomas, que actúa sobre las yodotirosinas MIT y DIT, pero no sobre T3 y T4. Casi todo el yoduro liberado se vuelve a usar para la síntesis de hormona, y normalmente sólo una pequeña cantidad esca- pa de la glándula tiroides (figura 7-10). Las 5′-desyodasa que convierte la T4 en T3 en los tejidos periféri- cos también se encuentra en la glándula tiroides. Cuando hay defi- ciencia de yoduro y en diversos estados hipertiroideos, la actividad de esta enzima aumenta la cantidad de T3 secretada por la glándu- la, lo que incrementa la eficiencia metabólica de la síntesis de hor- mona. ANORMALIDADES DE LA SÍNTESIS Y LIBERACIÓN DE HORMONA TIROIDEA Deficiencia de yodo en la dieta y defectos hereditarios Una dieta muy baja en yodo, y los defectos hereditarios en genes que codifican para las proteínas requeridas para la biosíntesis de hormona tiroidea (dishormonogénesis) pueden dar por resultado producción insuficiente de hormona. En respuesta a contenido de yodo o produc- ción de hormona, intratiroideo limitado, la glándula aumenta las http://booksmedicos.org 07Gardner(163-226).indd 168 07Gardner(163-226).indd 168 26/8/11 17:22:29 26/8/11 17:22:29
booksmedicos.org CAPÍTULO 7 Glándula tiroides 169 H H CH3 CH3 CH3CH2CH2 N S N N N S S S N COOC2H5 NH NH NH O O Carbimazol Tiouracilo Propiltiouracilo Metimazol FIGURA 7–9 Inhibidores tiocarbamida de la organificación de yoduro tiroidea. (TBG); la transtiretina, antes llamada prealbúmina de unión a tiroxi- na (TBP), y albúmina (figura 7-12). La unión a proteína plasmática permite el aporte de las yodotironinas mediante la sangre, que por lo demás son poco solubles en agua. También crea un fondo común grande de hormona tiroidea circulante, con una vida media plasmá- tica de siete días estable, y asegura la distribución homogénea de hormonas tiroideas en tejidos blanco. proporciones de MIT a DIT dentro de la tiroglobulina, así como la proporción de T3 secretada en comparación con T4. El eje hipo- tálamo-hipófisis-tiroideo también muestra respuesta a la deficiencia de hormona tiroidea al aumentar la secreción de TSH. En consecuen- cia, los individuos afectados típicamente se presentan con agranda- miento de la tiroides (bocio), que puede ser suficiente para compen- sar producción ineficiente de hormona tiroidea; sin embargo, de no ser así, presentan hipotiroidismo. Los recién nacidos y los lactantes gravemente afectados pueden sufrir los efectos irreversibles de la defi- ciencia de hormona tiroidea sobre el desarrollo que dan por resultado cretinismo. Los trastornos hereditarios específicos se describen con mayor detalle en la sección sobre bocio no tóxico, más adelante. Globulina de unión a tiroxina La globulina de unión a tiroxina (TBG) es un miembro glucoproteí- nico derivado del hígado, de la familia SERPIN de serina antiprotea- sas compuesta de una cadena polipeptídica de 54 kDa única, a la cual están fijas cuatro cadenas de carbohidrato que normalmente contie- nen alrededor de 10 residuos de ácido siálico. Cada molécula de TBG tiene un sitio de unión único para T4 o T3. La concentración sérica de TBG es de 15 a 30 μg/ml (280 a 560 nmol/L), y su afinidad de unión alta por la T4 y la T3 le permite transportar aproximadamente 70% de las hormonas tiroideas circulantes. La TBG y su unión de las hormonas tiroideas puede alterarse por trastornos congénitos de la TBG, ciertas circunstancias fisiológicas y fisiopatológicas, y varios fármacos. La deficiencia de TBG ocurre con una frecuencia de 1:5 000 nacidos vivos; se describen diversas varian- tes en varios grupos étnicos y raciales. Es un rasgo recesivo ligado a X que, en consecuencia, se expresa mucho más comúnmente en varo- nes. A pesar de concentraciones totales circulantes bajas de T4 y T3 en individuos afectados, las concentraciones de hormonas libres son normales, y estos pacientes permanecen eutiroideos. La deficiencia congénita de TBG a menudo se relaciona con deficiencia congénita de globulina de unión a corticosteroide (capítulo 9). Por el contrario, el exceso congénito de TBG, que es raro, se caracteriza por concen- traciones de T4 y T3 totales altas en sangre, pero cifras normales de hormonas libres y un estado clínico eutiroideo. El embarazo, los tu- mores secretores de estrógeno, y la terapia con estrógeno, aumentarán el contenido de ácido siálico de la molécula de TBG, lo que da por resultado decremento de la depuración metabólica y concentración sérica alta de TBG. Las cifras de TBG pueden disminuir en enferme- dades sistémicas importantes y, debido a división por proteasas de leucocitos, su afinidad de unión por las hormonas tiroideas también puede reducirse. Ambos efectos disminuyen las concentraciones de hormonas tiroideas totales séricas en pacientes enfermos (cuadro 7-1). Ciertos fármacos también pueden disminuir (esteroides androgéni- cos, glucocorticoides, danazol, l-asparaginasa) o aumentar (estróge- nos, 5-fluorouracilo) la concentración plasmática de TBG. Otros medicamentos (p. ej., salicilatos, fenitoína en dosis altas, y furosemida por vía intravenosa) pueden unirse a la TBG, lo cual desplaza la T4 y T3. En estas circunstancias, el eje hipotálamo-hipófisis-tiroideo (que se comenta más adelante) preserva concentraciones normales de hor- mona libre al disminuir las cifras séricas de hormona tiroidea total. Efectos del exceso de yodo sobre la biosíntesis de hormonas Aunque el yoduro es un sustrato esencial para la producción de hor- mona tiroidea, el exceso de yoduro en realidad inhibe tres pasos en la producción de hormona tiroidea: atrapamiento de yoduro, yodación de tiroglobulina (el efecto de Wolff-Chaikoff), y liberación de hor- mona desde la glándula. Estas acciones inhibidoras son transitorias, y la glándula tiroides normal escapa después de 10 a 14 días de estos efectos del exceso de yoduro. Estos efectos autorreguladores del yodu- ro aíslan la función tiroidea fisiológica de las fluctuaciones a corto plazo de la ingestión de yodo. Estas acciones del yodo excesivo también tienen implicaciones clínicas importantes; a veces causan disfunción tiroidea inducida por yodo y permiten el tratamiento con yoduro de ciertas enfermedades tiroideas. Si la tiroides es afectada por tiroiditis autoinmunitaria o ciertas formas hereditarias de dishormonogénesis, puede ser incapaz de escapar de la inhibición de la función glandular inducida por yo- duro, y es posible que aparezca hipotiroidismo. Por el contrario, una carga de yoduro puede inducir hipertiroidismo (efecto de Jod-Base- dow2) en algunos pacientes con un bocio multinodular, enfermedad de Graves latente, y rara vez en individuos con glándula tiroides que parece por lo demás normal. TRANSPORTE DE HORMONA TIROIDEA Ambas hormonas tiroideas circulan en la sangre unidas a proteínas plasmáticas; sólo 0.04% de la T4 y 0.4% de la T3 están no unidas o libres y, en consecuencia, disponibles para entrada a tejidos blanco y acción en los mismos (figura 7-11). Hay tres proteínas de transporte de hormona tiroidea principales: la globulina de unión a tiroxina 2Jod es la palabra alemana para yodo; Carl Adolph von Basedow fue uno de los pri- meros médicos en describir el hipertiroidismo. http://booksmedicos.org 07Gardner(163-226).indd 169 07Gardner(163-226).indd 169 26/8/11 17:22:29 26/8/11 17:22:29
booksmedicos.org 170 CAPÍTULO 7 Glándula tiroides 500 μg I Fondo común de yoduro (ECF) (150 μg I) 500 μg I 115 μg I 40 μg I 60 μg I Tracto GI Tejidos (hígado, músculo, etc.) Fondo común tiroideo (8 000 μg I) 15 μg I (bilis) 75 μg I (T4 + T3) 75 μg I (T4 + T3) Fondo común de hormona (T3 + T4) (600 μg I) 485 μg I (orina) 15 μg I (heces) FIGURA 7–10 Metabolismo del yodo. Los valores indicados son representativos de los que podrían encontrarse en un sujeto sano que ingiere 500 μg de yodo al día. La ingestión real de yodo varía considerablemente entre diferentes individuos. De modo similar, la estimulación con heparina de la lipoproteína li- pasa libera ácidos grasos libres que desplazan hormonas tiroideas de la TBG. In vivo, esto puede dar por resultado concentraciones más bajas de hormona tiroidea total, mientras que in vitro (p. ej., con sangre extraída a través de una llave de heparina), la heparina aumen- ta las concentraciones medidas de T4 y T3 libres. Control por retroacción Acción en los tejidos Secreción tiroidea T4 libre Metabolismo de hormona Transtiretina o prealbúmina fijadora de tiroxina (TBPA) La transtiretina, un polipéptido globular de 55 kDa, compuesto de cuatro subunidades de 127 aminoácidos idénticas, se une a 10% de la T4 circulante. Su afinidad por la T4 es 10 veces mayor que por la T3. La disociación de T4 y T3 desde la transtirretina es rápida, de modo que la transtiretina es una fuente de T4 fácilmente disponible. Como una enfermedad hereditaria, puede ocurrir aumento de la afinidad de la transtiretina por la unión a T4. Los afectados tienen T4 total alta pero T4 libre normal. Se ha reportado que la producción ectópica de transtiretina ocurre en pacientes con tumores pancreáticos y hepáti- cos, también causa hipertiroxinemia eutiroidea. Excreción fecal T4 unida a proteína (99.96%) FIGURA 7–11 Representación de la T4 libre (y de la T3 libre) como las hormonas biológicamente activas en el ámbito de la hipófisis y los tejidos periféricos. Casi todas las hormonas tiroideas circulantes en el plasma están unidas a proteína y carecen de actividad biológica. Este fondo común de hormona unida está en equilibrio con el fondo común de hormona libre. (Reproducida, con autorización, de DeGroot LJ, Stan- bury JB. The Thyroid and Its Diseases. 4th ed. Wiley; 1975.) Albúmina La albúmina se une a T4 y T3 con menor afinidad que la TBG o la transtiretina, pero su concentración plasmática alta da por resultado http://booksmedicos.org 07Gardner(163-226).indd 170 07Gardner(163-226).indd 170 31/8/11 20:24:31 31/8/11 20:24:31
booksmedicos.org CAPÍTULO 7 Glándula tiroides 171 CUADRO 7–1 Causas de cuantificaciones de tiroxina sérica con resultados anormales en individuos eutiroideos. Hipertiroxinemia eutiroidea ↑Unión a proteína plasmática ↑Globulina de unión a tiroxina (TBG) Hereditaria Efectos de estrógeno (embarazo, terapia con estrógeno) Hepatitis Fármacos: tamoxifeno, 5-fluorouracilo, clofibrato, metadona, heroína ↑Unión a transtiretina Hereditaria Producción paraneoplásica por tumores hepáticos y pancreáticos ↑ Unión a albúmina Hereditaria (hipertiroxinemia disalbuminémica familiar) ↓Conversión de T4 en T3a Enfermedad sistémica Medicaciones: amiodarona, agentes de contraste radiográficos, glucocorticoides, propranolol Terapia con tiroxina en el hipotiroidismoa Resistencia generalizada a la hormona tiroideaa Anticuerpo anti-T4 (interferencia con el ensayo) Hipotiroxinemia eutiroidea ↓Globulina de unión a tiroxina ↓Producción de TBG Hereditaria Andrógenos Fármacos: danazol, L-asparaginasa ↑ Depuración de TBG Síndrome nefrótico Enfermedad hepática grave Gastroenteropatía perdedora de proteína Enfermedad sistémicab Medicaciones Compuestos tiromiméticos exógenos (T3 [Cytomel])b Fenitoína y carbamazepinab Deficiencia de yodo (con T3 sérica normal)b Proteínas séricas − + γ β α2 α1 Albúmina 70 a 75 T4 % unida 15 a 20 5 a 10 − + TBG Albúmina TBPA 70 a 75 25 a 30 T3 % unida − + TBG Albúmina FIGURA 7–12 Representación esquemática de la distribución de T4 y T3 radiactivas entre proteínas de unión a hormona tiroidea séricas. Arriba: patrón electroforético en papel de proteínas séricas. En medio: se añadió T4 radiactiva al suero, y después quedó sujeta a electroforesis en papel. Los máximos representan la movilidad de la T4 radiactiva unida a diferentes proteínas séricas. (TBG, globulina de unión a hormona tiroidea; TBPA, prealbúmina de unión a tiroxina.) Abajo: se añadió T3 radiactiva al suero y se sujetó a electroforesis en papel. Los máximos indican la distribución relativa de T3 radiactiva unida a proteína. Los números arriba de cada máximo indican la distribución de hormona relativa entre las proteínas de unión en un adulto normal. (Reproducida, con autorización, de Rosenfield RL et al. Pediatric Nuclear Medicine. James AE Jr, Wagner HN Jr, Cooke RE, eds. Saunders; 1974.) a T4 tanto total como libre altas. b T4 tanto total como libre bajas. METABOLISMO DE LAS HORMONAS TIROIDEAS La glándula tiroides normal secreta alrededor de 100 nmol de T4 y sólo 5 nmol de T3 al día; se producen menos de 5 nmol de T3 reversa (rT3) metabólicamente inactiva (figura 7-13). La mayor parte del fondo común plasmático de T3 (80%) se deriva de monodesyodación de anillo externo periférico, o de 5′-monodesyodación, de T4 en teji- dos fuera de la glándula tiroides, en particular el hígado, los riñones y el músculo esquelético (cuadro 7-2). Dado que la T3 tiene afinidad de unión más alta por los receptores de T3 nucleares que afectan la acción de la hormona tiroidea, esta 5′-monodesyodación genera una yodotironina con mayor actividad biológica. Por otro lado, la desyo- dación del anillo interno de T4 (5-desyodación) produce 3,3′,5′- triyodotironina o T3 reversa (rT3), que es inerte desde el punto de vista metabólico. Las tres enzimas desyodasa que catalizan estas reacciones difieren en la localización en tejidos, la especificidad de sustrato y la modu- lación fisiológica y fisiopatológica, como se resume en el cuadro 7-2. La 5′-desyodasa tipo 1, la forma más abundante, se encuentra de manera predominante en el hígado y los riñones, y en menores can- tidades en la glándula tiroides, el músculo esquelético y cardiaco, y su transporte de 15% de T4 y T3 circulantes. Las tasas de disocia- ción rápida de hormona tiroidea desde la albúmina hacen de esta última una fuente importante de hormona libre para los tejidos. La hipoalbuminemia, como ocurre en la nefrosis o la cirrosis, se relacio- na con T4 y T3 totales bajas, pero las cifras de hormonas libres son normales. La hipertiroxinemia disalbuminémica familiar es un trastorno hereditario autosómico dominante en el cual 25% de la albúmina muestra una afinidad de unión a T4 más alta que lo normal. Esto da lugar a una concentración alta de T4 total pero cifras normales de T4 libre, y eutiroidismo. En casi todas las familias afectadas, la afinidad de unión a T3 es normal. Dado que estas variantes de albúmina no se unen a los análogos de tiroxina usados en muchos inmunoensayos de T4 libre, pueden reportar falsamente aumento de la T4 libre en los afectados. http://booksmedicos.org 07Gardner(163-226).indd 171 07Gardner(163-226).indd 171 26/8/11 17:22:29 26/8/11 17:22:29
booksmedicos.org 172 CAPÍTULO 7 Glándula tiroides celulares neuronales. Por el contrario, la T4 plasmática alta reduce la concentración de 5′-desyodasa tipo 2, lo que protege a las células del cerebro contra T3 excesiva. En consecuencia, esta desyodasa represen- ta un mecanismo mediante el cual el hipotálamo y la hipófisis pueden mostrar respuesta a la concentración de T4 circulante. La rT3 también puede modificar la actividad de la 5′-desyodasa tipo 2 en el cerebro y la hipófisis, y los compuestos α-adrenérgicos estimulan la 5′- desyodasa tipo 2 en la grasa parda, pero no está clara la importancia fisiológica de estos efectos. La 5-desyodasa tipo 3 se encuentra en las membranas coriónicas de la placenta y en las células gliales en el sis- tema nervioso central, donde desactiva la T4 al convertirla en rT3, y desactiva T3 al convertirla en 3,3′-diyodotironina (3,3′-T2) (figura 7-14). La concentración de desyodasa tipo 3 está alta en el hipertiroi- dismo y disminuida en el hipotiroidismo; por ende, puede ayudar a aislar al feto y el cerebro contra el exceso o la deficiencia de T4. La desyodasa tipo 3 placentaria acelera la eliminación de tiroxina en embarazadas, lo cual explica en parte los requerimientos de dosis au- mentadas de tiroxina en mujeres hipotiroideas tratadas. En general, las funciones de las desyodasas son importantes desde el punto de vista fisiológico de tres maneras. En primer lugar, permi- ten la modulación tisular y celular locales de las acciones de la hor- mona tiroidea. En segundo lugar, ayudan al organismo a adaptarse a estados cambiantes, incluso deficiencia de yodo o enfermedad cróni- ca. En tercer lugar, regulan las acciones de la hormona tiroidea en el desarrollo temprano de muchos vertebrados, incluso anfibios y ma- míferos. Alrededor de 80% de la T4 se metaboliza mediante desyodación, 35% hacia T3 y 45% hacia rT3 (figura 7-13). El resto se desactiva principalmente mediante glucuronidación en el hígado y secreción biliar, y en menor grado, mediante sulfatación en el hígado o los ri- ñones. Otras reacciones metabólicas incluyen desaminación de la ca- dena lateral alanina, lo que forma derivados del ácido tiroacético de actividad biológica baja, o descarboxilación o división del enlace éter, lo que forma compuestos inactivos. Como resultado de estas vías metabólicas, cada día se depura aproximadamente 10% del fondo común total de T4 extratiroidea de 1 000 nmol, y la vida media plasmática de la T4 es de siete días. De- bido a la afinidad de unión más baja de la T3 por las proteínas plas- máticas, el recambio de este fondo común extratiroideo de mucho menor tamaño es más rápido, con una vida media plasmática de un día. El fondo común corporal total de rT3 es de alrededor del mismo tamaño que el de T3, pero la rT3 tiene un recambio aún más rápido, con una vida media plasmática de sólo 0.2 días. <5 nmol 5 nmol 100 nmol/día 45 35 rT3 T4 T3 nmol nmol 20 nmol (TETRAC, etc.) FIGURA 7–13 Principales vías del metabolismo de la tiroxina en seres humanos adultos normales. Las tasas están expresadas en nmol/24 h, y son aproximaciones basadas en los datos disponibles. 100 nmol de T4 equivalen a aproximadamente 75 μg. (rT3, T3 reversa; TETRAC, ácido tetrayodotiroacético.) (Reproducida, con autorización, de Cavalieri RR, Rapoport B. Impaired peripheral conversion of thyroxi- ne to triiodothyronine. Ann Rev Med. 1977;28:5765.) otros tejidos. La 5′-desyodasa tipo 1 contiene una porción selenocis- teína que probablemente está en el sitio desyodante activo. La prin- cipal función de la 5′-desyodasa tipo 1 es proporcionar T3 a la circu- lación. Su actividad está aumentada en el hipertiroidismo y disminuida en el hipotiroidismo. Esto explica en parte las cifras de T3 circulantes relativamente más altas en comparación con la con- centración de T4 que se observa en pacientes hipertiroideos. La enzi- ma es inhibida por el fármaco antitiroideo tionamida PTU, pero no por el metimazol, y por el fármaco antiarrítmico amiodarona y colo- rantes de contraste radiactivo yodados, como el ipodato de sodio. La deficiencia de selenio en la dieta también puede alterar la conversión de T4 en T3. La 5′-desyodasa tipo 2 se expresa de modo predominante en el cerebro y la hipófisis, donde mantiene una concentración constante de T3 intracelular en el sistema nervioso central. Esta desyodasa es muy sensible a la T4 circulante, de modo que una T4 circulante más baja aumenta con rapidez la concentración de enzima en el cerebro y la hipófisis, al alterar la tasa de degradación y desactivación de T4. Al hacerlo, mantiene la concentración de T3 intracelular y sus funciones CUADRO 7–2 Tipos de yodotironina desyodasa, y características. Tipo de desyodasa D1 D2 D3 rT3> T4> T3 T4> rT3 T3> T4 Sustratos Distribución en tejido Hígado, riñones, músculo esquelético, tiroides Cerebro, hipófisis Cerebro, placenta, tejidos fetales Producción de T3 plasmática Producción local de T3 Degradación de T3 Función Inhibición por PTU (IC50, μM) >1 000 >1 000 5 Hipotiroidismo Disminución Aumento Disminución Hipertiroidismo Aumento Disminución Aumento http://booksmedicos.org 07Gardner(163-226).indd 172 07Gardner(163-226).indd 172 26/8/11 17:22:30 26/8/11 17:22:30
booksmedicos.org CAPÍTULO 7 Glándula tiroides 173 I I HO O R “Paso hacia abajo” I I “Paso hacia arriba” T4 I I I I HO O R HO O R I I T3 T3 reversa I I I I HO O R HO O R HO O R I I I I 3,3'-T2 3',5'-T2 3,5-T2 HO O R HO O R ? ? 3-T1 3'-T1 HO O R NH2 R = CH2 CH COOH T0 FIGURA 7–14 La vía desyodante del metabolismo de la tiroxina. La monodesyodación de T4 a T3 representa un paso hacia arriba en potencia biológica, mientras que la monodesyodación de T4 hacia T3 reversa tiene el efecto opuesto. La desyodación adicional de T3 en esencia suprime la actividad hormonal. CONTROL DE LA FUNCIÓN Y LA ACCIÓN HORMONAL TIROIDEAS El crecimiento y la función de la glándula tiroides están controlados por el eje hipotálamo-hipófisis-tiroideo (figura 7-15) y, como se co- mentó, por el yoduro mediante los elementos de autorregulación. La hormona liberadora de tirotropina (TRH) hipotalámica estimula las células tirotropas en la parte anterior de la hipófisis para que produz- can TSH que, a su vez, promueve el crecimiento de la glándula tiroi- des y la secreción de hormonas por la misma. Además, las desyodasas en la hipófisis y los tejidos periféricos modulan los efectos de la hor- mona tiroidea mediante su conversión específica para tejido, de T4 en la yodotironina más activa T3. Por último, los efectos moleculares de la T3 en tejidos individuales están modulados por el subtipo de receptor de T3 con el cual interactúa; la respuesta de activación o re- presión de gen específica que induce y, en un campo que apenas ahora se está revelando, la interacción de receptores de T3 con otros ligan- dos, receptores estrechamente relacionados (p. ej., receptor de retinoi- de X; RXR), y coactivadores y correpresores que interactúan con ella en la modulación de la expresión de gen. Hipotálamo TRH − T3 Sistema porta I Parte anterior de la hipófisis + T4 TSH “Libre” T3 − T3 T4 Tejido + I Hormona liberadora de tirotropina La hormona liberadora de tirotropina (TRH) es un tripéptido, piro- glutamil-histidil-prolina amida (piro-Glu-His-Pro-NH2), sintetizado por neuronas en los núcleos supraóptico y paraventricular del hipotá- lamo (figura 7-16). La TRH se almacena en la eminencia media del hipotálamo y después se transporta por medio del sistema venoso porta hipofisario en dirección descendente por el tallo hipofisario hacia la parte anterior de la hipófisis, donde controla la síntesis de TSH y la liberación de la misma. La TRH también se encuentra en Tiroides T4 FIGURA 7–15 El eje hipotálamo-hipófisis-tiroideo. La TRH produci- da en el hipotálamo llega a las células tirotropas en la parte anterior de la hipófisis mediante el sistema porta hipotálamo-hipofisario, y estimula la síntesis y liberación de TSH. Tanto en el hipotálamo como en la hipófisis, es principalmente T3 la que inhibe la secreción de TRH y TSH, respectivamente. La T4 pasa por monodesyodación hacia T3 en tejidos neurales e hipofisarios, así como periféricos. http://booksmedicos.org 07Gardner(163-226).indd 173 07Gardner(163-226).indd 173 26/8/11 17:22:30 26/8/11 17:22:30
booksmedicos.org 174 CAPÍTULO 7 Glándula tiroides respectivas. El receptor de TRH es un miembro de la familia de re- ceptor acoplado a proteína G, de siete dominios transmembrana (cuadro 1-1; figura 1-4). La TRH se une a la tercera hélice transmem- brana del receptor, y activa tanto su complejo productor de monofos- fato de guanosina cíclico como la cascada emisora de señales inositol 1,4,5-trifosfato (IP3) que libera Ca2+ intracelular y genera 1,2-diacil- glicerol, lo que activa la proteína cinasa C. Estas vías se encargan de estimular la liberación de TSH. Coordinan la transcripción de los genes que codifican para las subunidades de TSH y la glucosilación postraduccional de la TSH, que es necesaria para que tenga actividad biológica completa. La secreción de TSH estimulada por TRH es pulsátil (figura 7-17), con una amplitud de impulso de TSH media de 0.6 mU/L cada 2 h. Los individuos normales tienen un ritmo circadiano en la liberación de TSH, con una concentración circulante máxima entre la medianoche y las 4 a.m., que probablemente está controlada por un generador de impulsos neuronales hipotalámico que impulsa la síntesis de TRH. Las hormonas tiroideas ejercen retroacción negativa adicional sobre la producción de TSH en el ámbito de la hipófisis al regular en dirección descendente el número de receptores de TRH sobre células tirotropas hipofisarias. En consecuencia, en pacientes con hipertiroi- dismo, tanto los pulsos de TSH como su aumento repentino de se- creción nocturno están notoriamente suprimidos, mientras que en pacientes hipotiroideos la amplitud del pulso y el aumento nocturno N N H CH2 O O O O N NH2 C NH CH C C N H (piro)Glu-His-Pro-(NH2) FIGURA 7–16 Estructura química de la hormona liberadora de ti- rotropina (TRH). otras porciones del hipotálamo, el cerebro y la médula espinal, donde puede tener funciones separadas como un neurotransmisor. El gen pre-pro-TRH, que codifica para una molécula grande con cinco co- pias de la secuencia progenitora de TRH de Glu-His-Pro-Gli, está ubicado en el cromosoma 3. La expresión del gen TRH está regulada de manera negativa por la hormona tiroidea —tanto la T3 suminis- trada por la circulación como la que surge por desyodación de T4 en las neuronas peptidérgicas mismas (cuadro 7-2). En la parte anterior de la hipófisis, la TRH se une a un receptor de membrana específico situado en células secretoras de TSH y de prolactina, lo que estimula la síntesis y la liberación de sus hormonas 4 3 TSH (mU/ml) 2 1 0 0 240 480 720 960 1 200 1 440 Tiempo (minutos) 6 5 4 TSH (mU/ml) 3 2 1 0 0 240 480 720 960 1 200 1 440 Tiempo (minutos) = Periodos de sueño FIGURA 7–17 TSH sérica en dos sujetos normales que demuestra pulsos de secreción espontáneos y el ritmo circadiano de secreción de TSH. (El tiempo 0 es 0900; las estrellas indican pulsos importantes.) (Reproducida, con autorización, de Greenspan SL et al. Pulsatile secretion of TSH in man. J Clin Endocrinol Metab. 1986;63:664. Copyright 1986 por The Endocrine Society.) http://booksmedicos.org 07Gardner(163-226).indd 174 07Gardner(163-226).indd 174 26/8/11 17:22:30 26/8/11 17:22:30
booksmedicos.org CAPÍTULO 7 Glándula tiroides 175 Tirotropina (hormona estimulante de la tiroides) de la secreción de TSH son mucho mayores. En animales de experi- mentación y en seres humanos recién nacidos, la exposición a tempe- raturas frías aumenta la secreción de TRH y TSH. Otras hormonas y fármacos (p. ej., vasopresina y agonistas α-adrenérgicos) estimulan la síntesis de TRH y la liberación de la misma. La administración de TRH sintética por vía intravenosa a seres humanos como un bolo de 200 a 500 μg, genera una triplicación a quintuplicación expedita de las cifras séricas de TSH, que alcanzan un máximo a alrededor de los 30 min y dura 2 a 3 h (figura 4-14). En pacientes con hipotiroidismo primario, en quienes la TSH basal está alta, hay una respuesta de TSH exagerada a la TRH exógena, y esta respuesta está suprimida en pacientes con hipertiroidismo, terapia con tiroxina en dosis altas, e hipotiroidismo central. La TRH también se encuentra en las células de los islotes del páncreas, el tracto gastrointestinal, la placenta, el corazón, la próstata, los testículos y los ovarios. La T3 no inhibe la producción de TRH en estos tejidos periféricos. Aún se desconoce el papel de la TRH en estos tejidos. La hormona estimulante de la tiroides (TSH) es una glucoproteína de 28 kDa compuesta de subunidades α y β que están enlazadas de manera no covalente. La subunidad α es común a las otras dos glu- coproteínas hipofisarias, la hormona estimulante del folículo (FSH) y la hormona luteinizante (LH), y la hormona placentaria gonadotro- pina coriónica humana (hCG), mientras que la subunidad β es sin- gular para cada hormona glucoproteínica; confiere propiedades de unión y actividad biológica específicas. Los genes que codifican para las sub unidades α y β de la TSH están ubicados en los cromosomas 6 y 1, respectivamente. La subunidad α humana tiene un centro de apoproteína de 92 aminoácidos, y contiene dos cadenas de oligosacá- rido; la subunidad β de la TSH tiene un centro de apoproteína de 112 aminoácidos, y contiene una cadena de oligosacárido. Las cade- nas de aminoácidos de las subunidades α y β de la TSH forman, cada una, tres asas que están entremezcladas y forman un nudo cistina FIGURA 7–18 Configuración esquemática del complejo TSH-TSHR. La porción central de la figura representa la estructura parecida a cinta de la TSH dentro del receptor de TSH. La línea de color rosa representa la subunidad β, y la de color amarillo, la subunidad α. Véase también la figura 7-19. (Reproducida, con autorización, de Szkudlinski MW, et al. Thyroid-stimulating hormone and thyroid-stimulating hormone receptor structure-function relationships. Physiol Rev. 2002;82:473.) http://booksmedicos.org 07Gardner(163-226).indd 175 07Gardner(163-226).indd 175 26/8/11 17:22:30 26/8/11 17:22:30
booksmedicos.org 176 CAPÍTULO 7 Glándula tiroides Nueve repeticiones ricas en leucina (AA 58-277) Subunidad A c Hélice α Lámina β 1 Sitios de división Péptido C c c c c N-glucano c c c 2 c “Péptido inmunogénico” (AA 352-366) c c c NH7 c Subunidad B COOH704 FIGURA 7–19 Representación esquemática del receptor de TSH. La subunidad A es la porción de unión a ligando del receptor, y la subunidad B es la porción de activación. Los ligandos que se unen al receptor comprenden TSH, anticuerpo estimulante de TSH, y anticuerpo bloqueador de TSH. Hay dos sitios de división que permiten el rompimiento del receptor y la pérdida de la subunidad A hacia el suero. (Reproducida, con autorización, de Rapoport B et al. The thyrotropin (TSH)-receptor: interaction with TSH and autoantibodies. Endocr Rev. 1998;19:673.) ceptor de TSH pueden causar hipertiroidismo familiar, y mutaciones activadoras somáticas dan lugar a adenomas tóxicos. Otras mutacio- nes pueden llevar a activación aberrante de receptor de TSH por la hCG, la hormona glucoproteínica placentaria que es similar desde el punto de vista estructural a la TSH, en el hipertiroidismo gestacional familiar. (figura 7-18). La glucosilación tiene lugar en el RER y el aparato de Golgi, donde residuos de glucosa, manosa y fucosa, y residuos de sulfato y ácido siálico terminales se enlazan al centro de apoproteína. Tres residuos de carbohidrato prolongan su vida media plasmática y aumentan su capacidad para inducir activación de receptor de TSH. La TSH controla el crecimiento de células y la producción de hormonas de la tiroides, al unirse a un receptor de TSH específico (TSH-R) 1 de aproximadamente 1 000 ubicado en la membrana ce- lular basolateral de cada célula tiroidea. La unión a TSH activa las vías tanto del monofosfato de adenosina cíclico (cAMP) como del fosfo- inositol para la transducción de señal. El gen que codifica para el re- ceptor de TSH está ubicado en el cromosoma 14; el producto es una glucoproteína de una sola cadena, compuesta de 764 aminoácidos. El receptor de TSH es un miembro de la familia de receptores acoplados a proteína G, con siete dominios transmembrana, con un ectodomi- nio involucrado en la unión a ligando y una porción intramembrana e intracelular que se encarga de la activación de las vías emisoras de señales que promueven el crecimiento de células, y la síntesis y libe- ración de hormonas tiroideas (figura 7-19). El receptor de TSH está involucrado en la patogenia de muchas formas congénitas y adquiridas de hipotiroidismo e hipertiroidismo. Se han descrito defectos hereditarios que dan por resultado síntesis o acción alterada de la TSH, incluso genes mutantes que codifican para factores de transcripción requeridos para la diferenciación de células tirotropas hipofisarias (POU1F1, PROP1, LHX3, HESX1), receptor de TRH, cadena β de TSH, receptor de TSH, y Gsα, que transduce la unión de receptor de TSH hacia activación de adenilato ciclasa. Los anticuerpos bloqueadores de receptor de TSH adquiridos también pueden causar hipotiroidismo. El trastorno más común relacionado con receptor de TSH que causa hipertiroidismo es la enfermedad de Graves, en la cual autoan- ticuerpos se unen al receptor de TSH y lo estimulan. Empero, el re- ceptor de TSH está involucrado en la causa de varias otras formas de hipertiroidismo. Mutaciones de línea germinal que activan el re- Efectos de la TSH sobre la célula tiroidea La TSH tiene muchas acciones sobre la célula tiroidea. Casi todas sus acciones están mediadas por el sistema de proteína G-adenilil ciclasa- cAMP, pero también está involucrada la activación del sistema de fosfatidilinositol (PIP3) con un incremento resultante del calcio intra- celular. Las principales acciones de la TSH son: A. Cambios de la morfología de las células tiroideas La TSH induce con rapidez seudópodos en el borde célula folicular-coloide, lo que acelera la resorción de tiroglobulina. El contenido de coloide disminuye a medida que se forman gotitas de coloide intracelulares y se estimula la formación de lisosoma, lo que aumenta la hidrólisis de tiroglobulina y la liberación de hormona tiroidea. B. Crecimiento celular Células tiroideas individuales aumentan de tamaño; la vascularidad se incrementa y, al cabo de cierto periodo aparece agrandamiento de la tiroides, o bocio. C. Metabolismo del yodo La TSH estimula todas las fases del metabolismo del yodo, desde aumento de la captación y el transporte de yoduro hasta yodación aumentada de tiroglobulina e incremen- to de la secreción de hormonas tiroideas y de la tiroglobulina misma. La expresión aumentada de NIS y la estimulación de la producción de cAMP median transporte aumentado de yoduro, y la hidróli- sis de PIP3 y el incremento del Ca2+ intracelular estimulan la yoda- ción de tiroglobulina. El efecto de la TSH sobre el transporte de yo- http://booksmedicos.org 07Gardner(163-226).indd 176 07Gardner(163-226).indd 176 26/8/11 17:22:30 26/8/11 17:22:30
booksmedicos.org CAPÍTULO 7 Glándula tiroides 177 Los tumores y otros trastornos del hipotálamo o de la parte ante- rior de la hipófisis pueden alterar la secreción de TRH y TSH, respec- tivamente. Los trastornos hipofisarios que causan hipotiroidismo se llaman secundarios, mientras que la enfermedad hipotalámica que da por resultado hipotiroidismo se llama terciaria. El diagnóstico dife- rencial de estas lesiones se comenta más tarde (véase Pruebas tiroi- deas). duro a través de la célula es bifásico: al principio, está deprimido (flujo de yoduro hacia afuera) y después, luego de un retraso de varias horas, aumenta la captación de yoduro. El flujo de yoduro hacia afuera puede deberse al incremento rápido de la hidrólisis de la tiro- globulina, con liberación de hormona y escape de yoduro hacia afue- ra de la glándula. La estimulación de captación de yodo por los tiro- citos y secreción de tiroglobulina, mediada por TSH, también ocurre después de la administración de TSH recombinante para el trata- miento con yodo radiactivo y la vigilancia de pacientes con cánceres tiroideos bien diferenciados. Otros estimuladores e inhibidores de la tiroides El folículo tiroideo tiene un rico aporte de capilares que llevan fibras nerviosas noradrenérgicas desde el ganglio cervical superior, y fi- bras nerviosas acetilcolinesterasa positivas derivadas del nervio vago y de ganglios tiroideos. Las células C parafoliculares secretan tanto calcitonina como péptido relacionado con el gen de la calcitonina (CGRP). En animales de experimentación, estos neuropéptidos y otros modifican el flujo sanguíneo y la secreción de hormonas tiroi- deas. Además, factores de crecimiento como insulina, el factor de crecimiento insulínico tipo I (IGF-I), las proteínas de unión al factor de crecimiento tipo insulina, el factor de crecimiento epidérmico (EGF), factor de crecimiento transformante β1, y factores de creci- miento de fibroblastos y sus receptores, así como factores autocrinos como las prostaglandinas y las citocinas, modifican el crecimiento de células, y la producción de hormonas tiroideas. Aun así, todavía no está claro qué tan importantes en clínica son estos efectos. D. Otros efectos de la TSH Otros efectos son aumento de la transcripción de los mRNA que codifican para tiroglobulina y TPO; aumento de la incorporación de yoduro hacia MIT, DIT, T3 y T4, e incremento de la actividad lisosómica, con secreción aumentada de T4 y T3 desde la glándula. También hay actividad aumentada de la 5′-desyodasa tipo 1, que ayuda a conservar el yodo intratiroideo. La TSH tiene efectos adicionales sobre la glándula tiroides, inclu- so estimulación de la captación de glucosa, el consumo de oxígeno y la oxidación de glucosa. Hay recambio acelerado de fosfolípidos y es- timulación de la síntesis de precursores de purina y pirimidina, con síntesis aumentada de DNA y RNA. TSH sérica La TSH intacta y la subunidad α aislada están presentes en la sangre circulante, y son detectables mediante inmunoensayo en concentra- ciones que normalmente son de 0.5 a 4.0 mU/L y de 0.5 a 2 μg/L, respectivamente. La concentración sérica de TSH está aumentada en el hipotiroidismo primario y disminuida en la tirotoxicosis, sea endó- gena o por ingestión oral excesiva de hormonas tiroideas. La vida media plasmática de la TSH es de alrededor de 30 min, y la tasa de producción diaria es de aproximadamente 40 a 150 mU/día. La subunidad α glucoproteínica a menudo está desproporcionada- mente alta en pacientes con tumores hipofisarios secretores de TSH (véase más adelante); también está aumentada en mujeres posmeno- páusicas normales debido a incremento de la síntesis y secreción rela- cionado con aumento de la producción de gonadotropina. Función de las desyodasas hipofisarias y periféricas La 5′-desyodasa tipo 2 hipofisaria convierte la T4 en T3 en el cerebro y la hipófisis, y proporciona la principal fuente de T3 dentro de célu- las tirotropas. Su actividad se incrementa en el hipotiroidismo, e ini- cialmente mantiene una concentración intracelular normal de T3 en el cerebro a pesar de una concentración plasmática de T4 en disminu- ción. En el hipertiroidismo, un decremento de su actividad modera la exposición de células hipofisarias y neurales a exceso de hormona tiroidea. En contraste, la 5′-desyodasa tipo 1 está disminuida en el hipotiroidismo, lo que conserva T4, y aumentada en el hipertiroidis- mo, lo que acelera el metabolismo de T4 pero también lleva a incre- mento de la concentración sérica de T3 (cuadro 7-2). Control de la secreción hipofisaria de TSH Dos factores importantes controlan la síntesis y la liberación de TSH: 1) la concentración de T3 dentro de células tirotropas, que regula la expresión de mRNA, la traducción de TSH y la liberación de la hor- mona, y 2) TRH, que controla la glucosilación postraduccional y la liberación. La síntesis y liberación de TSH son inhibidas por concentraciones séricas altas de T4 y T3 (hipertiroidismo) y estimuladas por cifras bajas de hormona tiroidea (hipotiroidismo). Además, ciertas hormonas y fármacos inhiben la secreción de TSH; éstos comprenden somatosta- tina, dopamina, agonistas de la dopamina como bromocriptina, y dosis altas de glucocorticoides. La enfermedad grave puede causar inhibición de la secreción de TSH, y puede haber un aumento de rebote de la TSH a medida que el paciente se recupera. La magnitud de estos efectos varía; los fármacos mencionados por lo general sólo suprimen parcialmente la TSH sérica, pero aún es detectable. En contraste, el hipertiroidismo manifiesto puede suprimir la concentra- ción de TSH hasta por debajo de los límites de detección de incluso los inmunoensayos de TSH más sensibles. Autorregulación tiroidea La autorregulación puede definirse como la capacidad de la glándula tiroides para modificar su función a fin de adaptarse a cambios de la disponibilidad de yodo, independiente de la TSH hipofisaria. De este modo, los seres humanos pueden mantener secreción normal de hor- mona tiroidea con ingestiones de yoduro que varían de 50 μg a varios miligramos por día. Ya se comentaron algunos de los efectos de la deficiencia de yoduro o el exceso del mismo. La principal adaptación a la ingestión baja de yoduro es la síntesis preferencial de T3 más que de T4, lo que aumenta la eficacia metabólica de la hormona secretada. Por otro lado, el exceso de yoduro inhibe muchas funciones tiroideas, incluso el transporte de yoduro, la formación de cAMP, la generación de peróxido de hidrógeno, y la síntesis y secreción de hormonas. Al- gunos de estos efectos pueden estar mediados por la formación de ácidos grasos yodados intratiroideos. La capacidad de la tiroides nor- mal para escapar a estos efectos inhibidores (efecto de Wolff-Chaikoff) http://booksmedicos.org 07Gardner(163-226).indd 177 07Gardner(163-226).indd 177 26/8/11 17:22:31 26/8/11 17:22:31
booksmedicos.org 178 CAPÍTULO 7 Glándula tiroides Unión a DNA Unión a T3 1 5 2 1 2 0 4 1 0 COOH TR α1 + + + DNA NH2 1 5 2 1 2 0 370 4 9 0 − + COOH TR α2 DNA NH2 1 1 0 6 1 7 4 4 6 1 COOH + + + TR β1 NH2 DNA 1 1 5 9 2 2 7 5 1 4 COOH TR β2 + + + DNA NH2 FIGURA 7–20 Estructura proteínica deducida de los productos de gen α y β de receptor de hormona tiroidea. La proteína receptora tiene tres dominios: un dominio de unión a DNA con un alto grado de similitud entre los diferentes tipos de receptores, un dominio de unión a triyodotironina (T3) carboxilo terminal, y un dominio amino terminal que no se requiere para la función completa. Los números que aparecen arriba de las estructu- ras representan números de aminoácidos. A la derecha se muestran las propiedades de los receptores respecto a su capacidad para unirse a T3 y para unirse a un elemento de respuesta a T3 de DNA. El sombreado idéntico de dominios de receptor indica secuencias de aminoácidos idénticas. (TR, re- ceptor de hormona tiroidea.) (Reproducida, con autorización, de Brent GA. The molecular basis of thyroid hormone action. N Engl J Med. 1994;331:847.) permite a la glándula seguir secretando hormonas a pesar de una in- gestión alta de yoduro en la dieta. Es importante notar que esto difie- re del efecto terapéutico del yoduro en el tratamiento de enfermedad de Graves. Aquí, la concentración alta de yoduro inhibe de manera persistente en la endocitosis de tiroglobulina y la actividad lisosómica, lo que disminuye la liberación de hormona tiroidea y la concentra- ción de hormona circulante. Además, cantidades farmacológicas de yoduro reducen la vascularidad de la glándula, lo cual puede ser be- neficioso durante intervención quirúrgica de la tiroides por enferme- dad de Graves. De cualquier modo, este efecto también es transitorio; dura 10 a 14 días. un trastorno del movimiento en asociación con cifras séricas altas de T3 y bajas de T4. El receptor nuclear de T3 es un miembro de una superfamilia de receptores nucleares que incluye receptores nucleares para glucocorti- coides, mineralocorticoides, estrógenos, progestinas, vitamina D3 y retinoides (figura 1-16). Hay dos genes que codifican para receptor de hormona tiroidea (TR): TRα ubicado en el cromosoma 17, y TRβ en el cromosoma 3. Cada gen da al menos dos productos empalma- dos de manera diferencial, TRα 1 y 2, y TRβ 1 y 2, aunque se cree que el TRα 2 carece de actividad biológica (figura 7-20). Cada uno de estos receptores tiene un dominio de unión a ligando carboxilo terminal y un dominio de unión a DNA situado centralmente con dos dedos de cinc cisteína que facilitan su fijación específica a elemen- tos de respuesta a hormona tiroidea (TRE) en los promotores de genes blanco, y regulan su transcripción (figuras 1-16 y 1-17). La concentración tisular de estos receptores varía entre los tejidos y con su etapa de desarrollo; por ejemplo, el cerebro contiene de manera predominante TRα, el hígado en su mayor parte TRβ, y el músculo cardiaco contiene ambos. Se ha mostrado que mutaciones puntuales en el dominio de unión a ligando del gen TRβ son la causa del sín- drome de resistencia generalizada a hormona tiroidea (GRTH). Los TR se unen a TRE, que típicamente son secuencias de oligonucleóti- dos específicas, pareadas (p. ej., AGGTCA) (figura 7-21). Los TR también pueden funcionar como heterodímeros con receptores para otros factores de transcripción, como los receptores de retinoide X y de ácido retinoico. Los TRE en general están ubicados torrente arriba del sitio de inicio de la transcripción para las regiones codifica- doras de genes con capacidad de respuesta a hormona tiroidea. En genes regulados de manera positiva, los TR no unidos interactúan con correpresores (p. ej., correpresor de receptor nuclear [NCoR] y me- diador silenciador para receptores retinoico y de hormona tiroidea [SMRT]) para reprimir la transcripción basal al reclutar histona des- acetilasas que alteran la estructura de cromatina cercana. Cuando la T3 se une a TR estos complejos correpresores se liberan, y los TR unidos a T3 se asocian con complejos coactivadores que promueven la acetilación de histona local; también se asocian con otro complejo proteínico (proteína que interactúa con receptor de vitamina D/pro- Acciones de las hormonas tiroideas 1. LOS RECEPTORES DE HORMONA TIROIDEA Y MECANISMOS DE ACCIÓN Las hormonas tiroideas ejercen sus acciones por medio de dos meca- nismos generales: 1) acciones genómicas efectuadas mediante interac- ciones de T3 con sus receptores nucleares, lo que regula la actividad del gen, y 2) acciones no genómicas mediadas por interacciones de T3 y T4 con ciertas enzimas (p. ej., calcio ATPasa, adenilato ciclasa, piru- vato cinasa monomérica), transportadores de glucosa y proteínas mitocondriales. Las hormonas tiroideas, no unidas en el plasma, se transportan dentro de la célula, mediante transportadores específicos, entre ellos el transportador monocarboxilato 8 (MCT8), MCT10 y polipépti- do transportador de anión orgánico (OATP1C1). El OATP1C1 se expresa de manera predominante en los capilares del cerebro y en el plexo coroideo, y transporta T4 de preferencia, mientras que el MCT8 y el MCT10 se expresan en muchos tejidos y transportan tanto T4 como T3. Las hormonas tiroideas se transportan a través de la membrana celular hacia el citoplasma, y después hacia el núcleo, donde T3 se une a su receptor específico. A últimas fechas se han re- conocido síndromes clínicos relacionados con mutaciones en estos transportadores, como el síndrome de Allan-Herndon-Dudley en el cual los niños varones afectados presentan retraso mental, miopatía, y http://booksmedicos.org 07Gardner(163-226).indd 178 07Gardner(163-226).indd 178 26/8/11 17:22:31 26/8/11 17:22:31
booksmedicos.org CAPÍTULO 7 Glándula tiroides 179 nas tiroideas se unen a la proteína de membrana integrina αVβ3, que está involucrada en la activación (mediada por hormona tiroidea) de la cascada de MAP cinasa y estimulación de la angiogénesis. A Núcleo Correpresor 2. EFECTOS FISIOLÓGICOS DE LAS HORMONAS TIROIDEAS Los efectos transcripcionales de T3 demuestran de manera caracterís- tica un tiempo de retraso de horas a días para alcanzar el efecto com- pleto. Estas acciones genómicas tienen diversos efectos vitales, entre ellos crecimiento de tejido, maduración del cerebro, producción de calor y consumo de oxígeno, así como otros efectos específicos sobre el corazón, el hígado, los riñones, el músculo esquelético y la piel. Sin embargo, se cree que algunas acciones de la T3 no son genómicas, incluso su reducción de la actividad de 5′-desyodasa tipo 2 hipofisaria y el transporte aumentado de glucosa y aminoácidos que puede indu- cir en algunos tejidos. A continuación se resumen algunos efectos específicos de las hormonas tiroideas. TR-LBD TR-LBD Coactivador TR-DBD TR-DBD TRE Citoplasma B Correpresor Coactivador TR-LBD T3 RXR LBD TR LBD TR DBD RXR DBD TR DBD Transcripción T3 5'DI TRE Efectos sobre el desarrollo fetal La concentración de yoduro por el tejido tiroideo y la TSH hipofisa- ria aparece en el feto humano alrededor de las 11 semanas de gesta- ción. Debido al contenido placentario alto de la 5-desyodasa tipo 3, la mayor parte de la T3 y T4 maternas está desactivada, y muy poca hormona libre alcanza la circulación fetal. Sin embargo, esta pequeña cantidad de hormona libre proveniente de la madre puede tener im- portancia para el desarrollo temprano del cerebro fetal. Después de 15 a 18 semanas de gestación, el feto depende en gran parte de su propia secreción tiroidea. Aunque ocurre algo de crecimiento del feto en ausencia de secreción de hormona tiroidea fetal, el deterioro del desa- rrollo del cerebro y la maduración del esqueleto es notorio si no se diagnostica hipotiroidismo congénito y no se empieza terapia con hormona tiroidea con prontitud después del nacimiento. El hipoti- roidismo congénito da por resultado cretinismo, cuyas características comprenden retraso mental y enanismo. T4 T4 T3 T4 T3 TBP FIGURA 7–21 Modelo de la interacción de T3 con el receptor de T3. Panel A, fase inactiva: el dímero receptor de T3 no unido a ligando, unido al TRE junto con correpresores, actúa como un supresor de la transcripción de gen. Panel B, fase activa: T3 y T4 circulan unidas a pro- teínas de unión a tiroides (TBP). Las hormonas libres son transportadas hacia la célula mediante un sistema de transporte específico. Dentro del citoplasma, la 5’-desyodasa convierte la T4 en T3, y la T3 se mueve hacia el núcleo. Ahí se une al dominio de unión a ligando del monómero TR. Esto promueve la alteración del homodímero TR y heterodimerización con RXR sobre el TRE, desplazamiento de correceptores, y unión de co- activadores. El complejo TR-coactivador activa la transcripción de gen, lo que lleva a alteración de la síntesis de proteína y del fenotipo celular. (TR-LBD, dominio de unión a ligando del receptor de T3; TRDBD, domi- nio de unión a DNA de receptor de T3; RXR-LBD, dominio de unión a li- gando del receptor de retinoide X; RXR-DBD, dominio de unión a DNA de receptor de retinoide X; TRE, elemento de respuesta a hormona tiroi- dea; TBP, proteínas de unión a tiroxina; T3, triyodotironina; T4, tetrayodo- tironina, L-tiroxina; 5’DI, 5’-desyodasa.) Efectos sobre el consumo de oxígeno, la producción de calor y la formación de radicales libres La T3 aumenta el consumo de O2 y la producción de calor en parte mediante estimulación de la Na+-K+ ATPasa en todos los tejidos ex- cepto en el cerebro, el bazo y los testículos. Esto contribuye al índice metabólico basal (consumo de O2 somático total en reposo) aumen- tado, y el incremento de la sensibilidad al calor en el hipertiroidismo —y lo opuesto en el hipotiroidismo—. Las hormonas tiroideas es- timulan la mitocondriogénesis, lo que aumenta la capacidad oxidativa de la célula. También inducen cambios en la composición de proteína y líquido de la membrana interna mitocondrial que aumentan el metabolismo oxidativo mediante efectos tanto genómicos como no genómicos. La eficiencia reducida del metabolismo oxidativo causada por hormona tiroidea también se refleja en el aumento del paso por ciclos inútil de metabolitos intermediarios de carbohidratos. teína relacionada con TR) que recluta RNA polimerasa II y empieza la transcripción de gen. Algunos genes están regulados de manera negativa por TR unidos a T3, como los genes pre-pro-TRH y TSH subunidades α y β, pero en la actualidad se entienden menos bien los mecanismos moleculares involucrados. Las acciones de la hormona tiroidea para alterar niveles de expresión de mRNA específicos y sus proteínas producidas generan un conjunto de respuestas tisulares es- pecíficas (figura 7-21). Desde hace mucho tiempo se han sospechado acciones no genó- micas de las hormonas tiroideas con base en ciertas respuestas tisula- res rápidas que tienen lugar antes de que pudiera ocurrir transcripción de RNA, y por reconocimiento de sitios de unión a T3 y T4 fuera del núcleo. Por ejemplo, a últimas fechas se ha mostrado que las hormo- Efectos cardiovasculares (figura 7-22) La T3 estimula la transcripción de Ca2+ ATPasa del retículo sarcoplás- mico, lo que aumenta el índice de relajación diastólica miocárdica. http://booksmedicos.org 07Gardner(163-226).indd 179 07Gardner(163-226).indd 179 26/8/11 17:22:31 26/8/11 17:22:31
booksmedicos.org 180 CAPÍTULO 7 Glándula tiroides Triyodotironina Ca2+ Membrana celular Retículo sarcoplásmico Ca2+ Fosfolambán Núcleo Ca2+ ATPasa Ca2+ Elemento de respuesta a hormona tiroidea Ca2+ Receptor nuclear de triyodotironina Actina Miosina mRNA AMP cíclico Ca2+ Síntesis de proteína Proteína de unión a nucleótido guanina K+ Na+ Ca2+ Adenilil ciclasa K+ Receptor β-adrenérgico Na+/K+- ATPasa Na+ Canal de K+ sensible a voltaje Intercambiador de Na+/Ca2+ Triyodotironina FIGURA 7–22 Sitios de acción de la triyodotironina sobre miocitos cardiacos. La triyodotironina entra a la célula, posiblemente mediante un mecanismo de transporte específico, y se une a receptores de triyodotironina nucleares. El complejo a continuación se une a elementos de respuesta a hormona tiroidea de genes blanco, y regula la transcripción de estos genes. También ocurren efectos de hormona tiroidea no nucleares sobre ca- nales de iones. (Reproducida, con autorización, de Klein I, Ojamaa K. Thyroid hormone and the cardiovascular system. N Engl J Med. 2001;344:501.) Efectos simpáticos Las hormonas tiroideas aumentan el número de receptores β-adrenérgicos en el corazón y el músculo esquelético, el tejido adi- poso y los linfocitos. También pueden amplificar la acción de catecol- aminas en un sitio posreceptor. Muchas de las manifestaciones clíni- cas de la tirotoxicosis parecen reflejar sensibilidad aumentada a las catecolaminas. Además, la terapia con bloqueadores β-adrenérgicos a menudo es útil para controlar estas manifestaciones simpatomiméti- cas del exceso de hormona tiroidea. También aumenta la expresión de las isoformas más rápidamente contráctiles de la cadena pesada de miosina, las isoformas α, lo que contribuye a función sistólica aumentada. En el miocardio, la T3 también altera la expresión de diferentes isoformas de los genes que codifican para la Na+-K+ ATPasa, aumenta la expresión de receptores α-adrenérgicos, y disminuye la concentración de proteína G inhibi- dora Gi α. La T3 también aumenta los índices tanto de despolariza- ción como de repolarización del nodo sinoauricular, lo que aumenta la frecuencia cardiaca. En consecuencia, las hormonas tiroideas tienen efectos inotrópico y cronotrópico positivo sobre el corazón, lo cual, junto con el incremento de la sensibilidad adrenérgica (véase más adelante) explica la frecuencia cardiaca y la contractilidad aumentadas en el hipertiroidismo, y lo contrario en el hipotiroidismo. Las hormo- nas tiroideas también afectan la función ventricular diastólica, dismi- nuyen la resistencia vascular periférica y aumentan el volumen intra- vascular, lo cual contribuye más al incremento del gasto cardiaco relacionado con la acción de hormonas tiroideas. Efectos pulmonares Las hormonas tiroideas mantienen respuestas ventilatorias a la hi- poxia y la hipercapnia en el centro respiratorio del tallo encefálico. En consecuencia, en pacientes con hipotiroidismo grave, puede ocurrir hipoventilación. Las funciones de los músculos respiratorios también http://booksmedicos.org 07Gardner(163-226).indd 180 07Gardner(163-226).indd 180 26/8/11 17:22:31 26/8/11 17:22:31
booksmedicos.org CAPÍTULO 7 Glándula tiroides 181 están reguladas por la hormona tiroidea, y pueden estar debilitadas en el hipertiroidismo, lo que lleva a una sensación de falta de aliento. sis también está aumentada, con liberación de ácidos grasos y glicerol hacia el plasma circulante. Efectos hematopoyéticos Una demanda celular aumentada para O2 en el hipertiroidismo lleva a incremento de la producción de eritropoyetina y de la eritropoyesis. Sin embargo, por lo general no hay aumento del volumen sanguíneo debido a hemodilución y recambio aumentado de eritrocitos. Las hormonas tiroideas incrementan el contenido de 2,3-difosfoglicerato de los eritrocitos, lo que permite incremento de la disociación de O2 desde la hemoglobina, y aumenta la disponibilidad de O2 para los tejidos. Sucede lo contrario en el hipotiroidismo. Efectos endocrinos Las hormonas tiroideas alteran la producción, capacidad de respuesta y depuración metabólica de diversas hormonas. En niños hipotiroi- deos, la liberación alterada de hormona de crecimiento lentifica el crecimiento longitudinal. El hipotiroidismo puede causar retraso de la pubertad al alterar la secreción de hormona liberadora de gonadotro- pina (GnRH) y de gonadotropina. Por el contrario, el hipotiroidismo primario también puede causar pubertad precoz, quizá como un efec- to de concentraciones muy altas de TSH sobre receptores de gonado- tropina. En adultos, el hipotiroidismo causa hiperprolactinemia en una minoría de las mujeres afectadas. La menorragia (menstruaciones prolongadas y copiosas) y la anovulación son comunes en mujeres hi- potiroideas; esto último da lugar a esterilidad. La capacidad de respues- ta del eje hipotálamo-hipófisis-suprarrenal al estrés está aminorada en pacientes hipotiroideos. Una lentificación de la tasa de depuración metabólica de cortisol compensa para esto en el estado hipotiroideo. Empero, por el contrario, la restitución del eutiroidismo rara vez puede suscitar insuficiencia suprarrenal puesto que el metabolismo del cortisol está acelerado en pacientes con reserva de cortisol disminuida debido a enfermedad concomitante que afecta el eje suprarrenal. En el hipertiroidismo, la aromatización acelerada de andrógenos hacia estrógenos y el aumento de la concentración de globulina trans- portadora de hormonas sexuales contribuyen a la ginecomastia y a la concentración total alta de testosterona que se observan en los varones afectados. El hipertiroidismo también puede alterar la regulación nor- mal de la ovulación y las menstruaciones por la GnRH y la gonadotro- pina, lo que causa esterilidad y amenorrea, respectivamente. Todas las alteraciones endocrinas que ocurren con disfunción tiroidea se revier- ten mediante tratamiento apropiado para restituir el eutiroidismo. Efectos gastrointestinales Las hormonas tiroideas promueven la motilidad intestinal, lo cual puede dar lugar a incremento de la motilidad e hiperdefecación (esto es, frecuencia aumentada de defecaciones con heces formes) en el hipertiroidismo. Por el contrario, en el hipotiroidismo se observan tránsito intestinal lentificado y estreñimiento. Efectos esqueléticos Las hormonas tiroideas estimulan el recambio óseo, lo que aumenta la resorción ósea y, en menor grado, la formación de hueso. En consecuen- cia, el hipertiroidismo puede relacionarse con hipercalciuria, y menos a menudo, con hipercalcemia. Además, el exceso crónico de hormona tiroidea puede causar pérdida de mineral óseo importante en clínica. Efectos neuromusculares En el hipertiroidismo hay aumento del recambio de proteína y pérdi- da de músculo esquelético, lo cual puede llevar a una miopatía proxi- mal característica. Asimismo, hay un aumento de la rapidez de la contracción y relajación musculares, que se nota en clínica como la hiperreflexia propia del hipertiroidismo y la fase de relajación retrasa- da de reflejos tendinosos profundos en el hipotiroidismo. Un temblor distal fino de las manos también es típico del hipertiroidismo. Como se mencionó, las hormonas tiroideas son esenciales para el desarrollo y la función normales del sistema nervioso central, y el fracaso de la función tiroidea fetal da por resultado retraso mental grave. En el adulto, la hiperactividad en el hipertiroidismo y el aletargamiento en el hipotiroidismo pueden ser notorios. De modo similar, los pacientes hipertiroideos pueden estar ansiosos o agitados, mientras que el hipo- tiroidismo típicamente se relaciona con estado de ánimo deprimido. CAMBIOS FISIOLÓGICOS EN LA FUNCIÓN TIROIDEA Función tiroidea en el feto Antes de que el feto desarrolle su propia función tiroidea indepen- diente, su desarrollo neural temprano depende de las hormonas tiroi- deas maternas. Con todo, hacia la undécima semana de gestación, el sistema porta hipofisario se ha desarrollado, y hay TSH y TRH me- dibles. Casi al mismo tiempo, la tiroides fetal empieza a atrapar yodo. La secreción de hormona tiroidea probablemente empieza a la mitad de la gestación (18 a 20 semanas). La TSH aumenta con rapidez hasta concentración máxima a las 24 a 28 semanas, y las cifras de T4 alcan- zan un máximo a las 35 a 40 semanas. La concentración de T3 per- manece baja durante la gestación; la 5′-desyodasa tipo 3 convierte la T4 en rT3 durante el desarrollo fetal. En el momento del nacimiento, hay un aumento notorio repentino de la TSH estimulado por la ex- posición al ambiente extrauterino más frío, con incremento de T4 y de T3, y disminución de rT3; estos parámetros se estabilizan de ma- nera gradual en el transcurso del primer mes de vida. Efectos sobre el metabolismo de lípidos y carbohidratos El hipertiroidismo aumenta la gluconeogénesis y la glucogenólisis hepáticas, así como la absorción intestinal de glucosa, y puede haber también decrementos de la sensibilidad a la insulina mediados por hormona tiroidea. De este modo, el hipertiroidismo puede empeorar el control de la glucemia en pacientes que también tienen diabetes mellitus. Las hormonas tiroideas aumentan tanto la síntesis de coles- terol como la degradación del mismo. Este último efecto se debe en su mayor parte a un aumento del número de receptores de lipopro- teínas de baja densidad (LDL) hepáticos, lo que acelera la depuración de LDL. En consecuencia, las concentraciones de colesterol total y de LDL típicamente están altas en sujetos con hipotiroidismo. La lipóli- Función tiroidea durante el embarazo Varios parámetros tiroideos cambian durante la gestación. Hay un aumento de la depuración urinaria de yoduro; en áreas de ingestión http://booksmedicos.org 07Gardner(163-226).indd 181 07Gardner(163-226).indd 181 26/8/11 17:22:32 26/8/11 17:22:32
booksmedicos.org 182 CAPÍTULO 7 Glándula tiroides CUADRO 7–3 Condiciones o factores relacionados con decremento de la conversión de T4 o T3. baja de yodo en la dieta esto puede causar bocio materno o, cuando es grave, hipotiroidismo. Durante el embarazo hay un aumento de la TBG debido a un incremento de la glucosilación hepática de TBG inducido por estrógeno, lo que prolonga su tasa de depuración meta- bólica. En consecuencia, la concentración sérica de T4 total y, en menor grado, la de T3 total, aumentan. Las cifras crecientes de hCG, que tiene una actividad agonista débil de receptor de TSH, contribu- yen a agrandamiento tiroideo mínimo. La hCG materna alcanza un máximo alrededor de las 12 semanas, lo que da lugar a concentración de T4 libre en el suero normal alta o incluso modestamente alta de forma trasitoria y supresión fisiológica de la TSH sérica hasta el rango normal bajo o incluso subnormal. La concentración patológicamente alta de hCG que se encuentra en mujeres con embarazo molar o co- riocarcinoma puede causar hipertiroidismo manifiesto. Las mujeres con vómito grave durante el embarazo, llamado hiperemesis gravídi- ca, también pueden tener concentración más alta de hCG, y presentar una tirotoxicosis gestacional transitoria. Como se mencionó, la des- yodasa tipo 3 placentaria acelera la depuración de tiroxina durante la gestación, lo cual explica en parte el requerimiento aumentado de dosis de tiroxina en mujeres hipotiroideas tratadas, y ocasionalmente causa hipotiroidismo leve en mujeres con tiroiditis autoinmune y re- serva disminuida de la glándula tiroides. El suministro transplacentario de sustancias relacionadas con la tiroides puede tener consecuencias sobre el feto. El yoduro materno que cruza la placenta es esencial para la producción de hormona tiroi- dea fetal; de cualquier modo, las cantidades grandes de yoduro en realidad pueden inhibir la función tiroidea y causar un bocio consi- derable, suficiente para obstruir el paso por el canal del parto. En mujeres con enfermedad tiroidea autoinmunitaria, las inmunoglobu- linas maternas que estimulan el receptor de TSH o lo bloquean pue- den cruzar la placenta y causar disfunción tiroidea en el feto. Los fármacos antitiroideos metimazol y PTU cruzan la placenta, y en dosis grandes alteran la función tiroidea fetal (capítulo 16). 1. Vida fetal 2. Restricción calórica 3. Enfermedad hepática 4. Enfermedad sistémica mayor 5. Fármacos: Propiltiouracilo Glucocorticoides Propranolol (efecto leve) Agentes de contraste radiográficos yodados (ácido iopanoico, ipodato de sodio) Amiodarona 6. Deficiencia de selenio to. A continuación la tasa de recambio de T4 es estable hasta después de los 60 años de edad, cuando de nuevo empieza a declinar. En an- cianos, la depuración metabólica de T4 puede disminuir hasta 50%, lo que exige una reducción de la dosis de T4 para pacientes tratados. Efectos de la enfermedad aguda y crónica sobre la función tiroidea (síndrome del eutiroideo enfermo) La enfermedad aguda o crónica tiene varios efectos notorios sobre la función tiroidea y la economía de la hormona. El efecto más común y más temprano es la inhibición de la conversión de T4 en T3, con un decremento resultante de la concentración circulante de T3. Este cam- bio se acompaña de un aumento de la concentración sérica de T3 re- versa, debido tanto a su conversión disminuida en 3,3′-T2 como a algo de aceleración de su producción a partir de T4. Estos cambios ocu- rren de manera fisiológica en el feto, y patológica en circunstancias de restricción calórica, como en la malnutrición, inanición y anorexia nerviosa, y en pacientes con diversas enfermedades sistémicas agudas o crónicas (cuadro 7-3). Se cree que la patogenia de este síndrome de T3 baja comprende citocinas, como el factor de necrosis tumoral secretado por células inflamatorias que inhibe la 5′-desyodasa tipo 1. Hay fárma- cos que también pueden inhibir la 5′-desyodasa tipo 1 y disminuir la concentración sérica de T3 en pacientes tratados; estos medicamen- Cambios de la función tiroidea con el envejecimiento El recambio de T4 es más alto en lactantes y niños, y después de la pubertad disminuye de modo gradual hasta las cifras propias del adul- T3-T4 baja Eutiroideo T3 baja + + rT3 FT4 TT4 Rango normal Rango normal T3 Mortalidad − − Gravedad de la enfermedad FIGURA 7–23 Representación esquemática de los cambios de los valores de hormona tiroidea séricos con la gravedad creciente de enfermedad no tiroidea. La disminución de los valores de T4 total (TT4) sérica y de T4 libre (FT4) se acompaña de una tasa de mortalidad en aumento rápido. (Re- producida, con autorización, de Nicoloff JT. Abnormal measurements in nonendocrine illness. En: Medicine for the Practicing Physician. 2nd ed. Hurst JW, ed. Butterworth-Heinemann; 1991.) http://booksmedicos.org 07Gardner(163-226).indd 182 07Gardner(163-226).indd 182 26/8/11 17:22:32 26/8/11 17:22:32
booksmedicos.org CAPÍTULO 7 Glándula tiroides 183 tos comprenden corticosteroides, amiodarona, colorantes colecistográ- ficos yodados, la PTU y propranolol en dosis altas. Al principio, la concentración sérica baja de T3 típicamente se acompaña de cifras de T4 total y libre normales o, menos comúnmente, un poco altas (figura 7-23). Sin embargo, a medida que la enfermedad se hace más grave, también hay una declinación de la T4 total y libre sérica. Esta hipoti- roxinemia puede tener varias causas. En ciertas enfermedades (p. ej., nefrosis y enfermedad grave del hígado), puede haber una declinación de la concentración circulante de TBG. Un aumento de los ácidos grasos libres plasmáticos puede interferir con la unión de T4 a la TBG, lo que libera más T4 libre que, a su vez, ejerce una retroalimentación sobre el hipotálamo y la hipófisis para reducir la TSH y establecer un nuevo equilibrio con una T4 total más baja. La TSH también puede quedar suprimida por citocinas circulantes en pacientes muy graves, y por la dopamina o los corticosteroides a menudo usados para tratar pacientes muy graves. Por último, la depuración de T4 está acelerada en la enfermedad. Todos estos factores pueden contribuir a las concentra- ciones séricas más bajas de T4 y de T4 libre, según se mide mediante los inmunoensayos de hormona libre, y se estima mediante procedimien- tos de captación de T3 en resina. No obstante, se ha reportado que la concentración de T4 libre medida mediante diálisis de equilibrio es normal en la mayoría de los pacientes, mas no en todos. Esto sugiere que la concentración sérica baja de T4 libre reportada por casi todos los ensayos de T4 libre disponibles en el comercio puede ser un artefacto de medición que es singular para pacientes muy graves. Surgen dos preguntas clínicas en pacientes enfermos que tienen este conjunto de anormalidades en las pruebas de función tiroidea. En primer lugar, puede ser desafiante distinguir entre el síndrome del eutiroideo enfermo e hipotiroidismo central; en ambas enfermedades, las concentraciones de T4 y TSH pueden estar bajas. La información clínica puede ser útil, incluso un antecedente de enfermedad hipofisaria o tiroidea conocida, traumatismo o intervención quirúrgica del encéfa- lo, o manifestaciones de otros elementos de hipopituitarismo (p. ej., testículos pequeños en un varón). Las pruebas de laboratorio adiciona- les a veces pueden brindar información. La concentración sérica de T4 libre cuantificada mediante diálisis de equilibrio típicamente es normal en el síndrome del eutiroideo enfermo. En mujeres posmenopáusicas, una FSH sérica alta atestigua al menos una función hipofisaria intacta, aunque la concentración de gonadotropina también puede estar baja en presencia de enfermedad grave. La medición de cortisol sérico puede ser en particular útil, porque debe ser normal o estar alta en cualquier paciente con enfermedad sistémica importante. Si es normal o está alta, es menos probable la enfermedad hipotalámica o hipofisaria. Sin em- bargo, a veces es necesario obtener estudios de imágenes urgentes del hipotálamo o la hipófisis para resolver la cuestión. Estas anormalidades en las pruebas de función tiroidea se normalizan cuando el paciente se recupera. Empero, la recuperación se puede acompañar de aumento de rebote de la TSH sérica que puede malinterpretarse como hipotiroidis- mo. En estas circunstancias, en ausencia de hipotiroidismo manifiesto en clínica, es mejor evitar la terapia con hormona tiroidea, y repetir la evaluación algunas semanas después de la recuperación. La segunda pregunta clínica es si el síndrome del eutiroideo enfer- mo en sí debe tratarse con hormona tiroidea. Ciertamente, muchos pacientes muy graves tienen problemas clínicos que recuerdan hipo- tiroidismo, como hipotermia, sensorio alterado, disfunción ventilato- ria y miocárdica, e hiponatremia. Se ha postulado que en estos pa- cientes puede haber hipotiroidismo intracelular, y que la terapia con hormona tiroidea podría ser beneficiosa. Por el contrario, se ha pro- puesto que estos cambios del estado tiroideo de hecho son mecanis- mos compensadores para reducir las tasas de consumo de oxígeno y de catabolismo de proteína, y que la terapia con hormona tiroidea en realidad podría ser perjudicial. Más aún, en dos estudios con asigna- ción al azar, en series clínicas pequeñas, y en modelos de enfermedad sistémica en animales, no se ha logrado demostrar beneficio alguno de la terapia con hormona tiroidea en estas circunstancias. Aunque el tema persiste sin resolverse, la práctica actual en general es no dar terapia con hormona tiroidea a menos que haya evidencia clínica y bioquímica convincente de hipotiroidismo verdadero. AUTOINMUNIDAD TIROIDEA La autoinmunidad está involucrada en la patogenia de muchas enfer- medades tiroideas, entre ellas enfermedad de Graves hipertiroidea, hipotiroidismo con tiroiditis autoinmunitaria o de Hashimoto, tiroi- ditis silenciosa y posparto, y ciertas formas de disfunción tiroidea neonatal. La defensa inmunitaria contra sustancias extrañas y células neoplásicas comprende macrófagos que ingieren y digieren el material extraño y presentan fragmentos de péptido sobre la superficie celular en relación con una proteína clase II codificada por la región HLA- DR del complejo de gen MHC. Este complejo es reconocido por un receptor de célula T sobre una célula T auxiliar CD4, que estimula la liberación de citocinas como interleucina-2 (IL-2). Estas citocinas amplifican la respuesta al inducir la activación y proliferación de cé- lulas T, la inducción de actividad de células asesinas en células supre- soras CD8, y la estimulación de la formación de anticuerpos contra el antígeno extraño por linfocitos B. Finalmente, el proceso de activa- ción es silenciado por la acción de las células supresoras CD8 (capí- tulo 2). Observaciones tempranas sugieren la posibilidad de que las reac- ciones inmunitarias podrían afectar la glándula tiroides. Se indujo tiroiditis mediante inmunización de ratones con tejido tiroideo y adyuvante de Freund. Se identificaron anticuerpos contra tiroglobu- lina en el suero de pacientes con tiroiditis, y se encontró un estimu- lador tiroideo de acción prolongada, que más tarde se mostró que es un anticuerpo contra el receptor de TSH, en el suero de pacientes con enfermedad de Graves. Hay tres autoantígenos tiroideos principales: tiroglobulina, TPO y el receptor de TSH. Los autoanticuerpos circulantes contra estos antígenos son marcadores útiles para autoinmunidad tiroidea, pero los mecanismos inmunitarios mediados por células T son fundamen- tales para la patogenia de enfermedades tiroideas. Las células tiroideas tienen la capacidad de ingerir antígeno (p. ej., tiroglobulina) y cuan- do son estimuladas por citocinas como el interferón gamma, expresan moléculas clase II de superficie celular (p. ej., HLA-DR4) que presen- tan estos antígenos a linfocitos T. Además del antígeno y la molécula clase II, para el inicio de enfermedad tiroidea autoinmunitaria sin duda se requieren otros factores, como señales coestimuladoras pro- venientes de células presentadoras de antígeno. Una hipótesis es que el proceso es iniciado o promovido por antígenos externos (p. ej., agentes infecciosos) que llevan a respuestas inmunitarias de anticuer- pos y celulares por medio de reactividad cruzada. Además, los factores genéticos claramente predisponen a individuos a autoinmunidad ti- roidea. La tipificación de HLA en pacientes con enfermedad de Graves revela una frecuencia alta de HLA-B8 y HLA-DR3 en caucá- sicos, HLA-Bw46 y HLA-B5 en chinos, y HLA-B17 en sujetos de http://booksmedicos.org 07Gardner(163-226).indd 183 07Gardner(163-226).indd 183 26/8/11 17:22:33 26/8/11 17:22:33