Download
1 / 86
860 likes | 1.46k Views
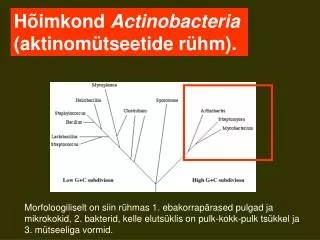
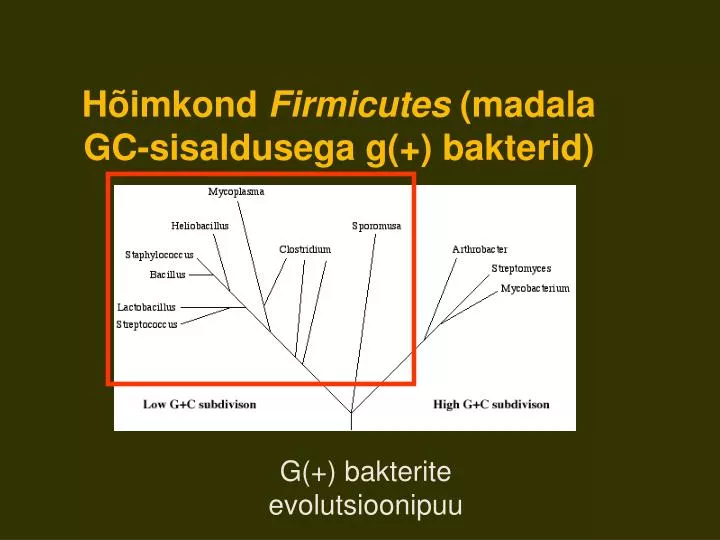
Hõimkond Firmicutes (madala GC-sisaldusega g(+) bakterid). G(+) bakterite evolutsioonipuu. Eristatakse 3 klassi: Klostriidid, Mollicutes e. mükoplasmad ja Batsillid.

E N D
Hõimkond Firmicutes (madala GC-sisaldusega g(+) bakterid) G(+) bakterite evolutsioonipuu
Eristatakse 3 klassi: • Klostriidid, • Mollicutes e. mükoplasmad ja • Batsillid. • Neis rühmades on erineva rakukujugavorme ja nii sporogeene kui ka asporogeene. rRNA järjestused näitavad, et mükoplasmad on lähedased klostriididega.
Saab rühmadesse jagada selle järgi, kas rakukest on olemas ja kas endospoore moodustatatakse.
Erinevatel g(+) bakterite rühmadel on erinev peptodoglükaani ehitus. Näiteks perekondades Bacillus, Clostridium, Lactobacillus, Corynebacterium, Mycobacterium ja Nocardia, on tetrapeptiidid ühendatud nii, nagu g(-) bakteritel: sideme moodustavad kolmandas positsioonis asuva DAP aminorühm ja naaberahela terminaalse D-Ala karboksüülrühm. Teistel G(+) bakteritel aga on DAP asemel Lys ja tetrapeptiidid liituvad lisapeptiidi kaudu. Need lisapeptiidid koosnevad monokarboksüülaminohapetest või glütsiinist või mõlemast. Selline peptidoglükaan on stafülokokkidel, streptokokkidel, mikrokokkidel, laktobatsillidel ja leukonostokitel.
Pentaglütsiinsild Staphylococcus aureuse peptidoglükaanis on tetrapeptiidid seostunud omavahel pentaglütsiinsillaga. Selline lisapeptiidiga seostumine on iseloomulik g(+) bakterite peptidoglükaanile
Huvitav peptidoglükaan on taimedele patogeensetel korünebakteritel. Neil moodustub side ühe ahela teises ja teise ahela neljandas asendis oleva tetrapeptiidse AH vahel ja kuna neil aminohapetel on sideme moodustamiseks vabad vaid karboksüülrühmad, siis on lisapeptiid moodustunud diaminohapetest, näiteks ornitiinist.
Klass Klostriidid (“Clostridia”) Suhteliselt kirju rühm. Siia kuulub ka nn ebatavalise rakukestaga baktereid, mis võivad värvuda gramnegatiivselt, aga on tegelikult tüüpilise g(+) ehitusega peptidoglükaaniga. Peptidoglükaankihte ilmselt on neil lihtsalt vähem. Clostridium botulinum
Klass Klostriidid (“Clostridia”). Klassi kuuluvad näiteks perekonnad: Clostridium, Sarcina, Butyrivibrio, Lachnospira, Ruminococcus, Eubacterium, Peptococcus, Desulfotomaculum, Propionispira, heliobakterid, Pectinatus, Megasphaera, Veillonella, Haloanaerobacter.
Perekonnas Clostridium on 146 liiki. Suhteliselt heterogeenne. Võimalik, et tehakse sellest perekonnast perekondi juurde. Siia kuuluvad obligaatselt anaeroobsed pulkbakterid, kes moodustavad termoresistentseid endospoore ja sulfaate ei redutseeri. Siia kuulub rida ohtlikke baktereid. C. botulinum põhjustab toidumürgitust botulismi, C. tetanikangestuskramptõbe, C. perfringens ja C. histolyticumgaasgangreeni ja ka toidumürgitust. C. difficile.Avastati 1930. a., kuid 70.ndatel hakati teda rohkem uurima seoses AB ravi järgse kõhulahtisusega. Algul peeti normaalse mikrofloora kahjutuks esindajaks, enam mitte. Põhjustab soolte põletikulist haavandumist. Klostriidid on ka biotehnoloogias olulised, näiteks saab nende abil atsetooni ja butanooli toota. Klostriididel pole tsütokroome ja neil töötab seetõttu ainult substraatne fosforüülimine. Substraadid, mida klostriidid kääritavad, on väga erinevad, samuti ka produktid, mis tekivad.
Sarcina. Anaeroobid. Kuubid pakendites (8 ja enam rakku). Taluvad väga hästi happelist keskkonda. Rakukest sisaldab tselluloosi ja see liidab ka agregaate. Sarcina liike saab isoleerida mullast, mudast, väljaheidetest, maosisaldistest. S. ventriculi (pildil) elab inimese alahappelises maos. Leitud just haavanditega patsientidel. Peptostreptococcus. Obligaatsed anaeroobid. Kääritavad aminohappeid, suhkruid mitte. Prpteolüütiliselt väga aktiivsed. Kuuluvad inimese normaalsesse mikrofloorasse (nahk, jämesool, tupp). On ühed tüüpilised anaeroobsete infektsioonide põhjustajad. Esinevad sageli koos Porfüromonase, Prevotella ja Bacteroidesega. P. magnus on põhiline põletikke tekitav liik. Põhjustab sünnitusjärgset emakapõletikku, põskkoopapõletikke, kroonilist keskkõrvapõletikku jt põletikke.
Eubacterium. Anaeroobsed jämesoolebakterid. Osa proteolüütilised, osa sahharolüütilised. Butyrivibrio. Võihapekääritaja. Leidub näiteks mäletsejate loomade vatsas. Lachnospira. Võihapekääritaja. Vatsabakter. Ruminococcus.Moodustab diplokokke ja ahelaid. Obligaatne anaeroob. Elab vatsas, jämesooles. Acidaminococcus. Range anaeroob. Kääritab aminohappeid. Desulfotomaculum. Sufaate redutseeriv sporogeenne bakter. Värvub gramnegatiivselt (peptidoglükaani vähem), kuid rakukesta ehitustüüp on g(+).
Eubacteriumon saledate rakkudega asporogeensed anaeroobid, keda leidub rohkesti inimese jämesooles, aga ka suuõõnes.
Heliobakterid On fotosünteesivad G(+) bakterid. Värvuvad gramnegatiivselt, kuid rakukesta ehitustüüp on g(+). Väga tundlikud penitsilliinile. Peptidoglükaankiht on lihtsalt õhem ja sellest g(-) värvumine. Neil on ebatavaline bakteriklorofüllBchlg,mille esinemine eristab heliobaktereid teistest fotosünteesivatest bakteritest, rohelistest ja punastest, kellel on teised klorofüllid ja Bchl g-d ei ole. Ka ei ole neil klorosoome ja FS pigmendid paiknevad neil TPMs.
Heliobacillus Heliobakteritel eristatakse 4 perekonda: Heliobacterium, Heliobacillus, Heliophilum ja Heliorestis. Levinud troopilises mullas (riisipõllud) Heliobacterium chlorum on pulgakujuline libisevalt liikuv, Heliobacterium gestii on spiraalne, liikuv, Heliobacterium fasciculum ja Heliobacillus mobilis on pulgakujulised. Heliophilum on pulkbakter, mis moodustab koos liikuvaid rakkude kimpe. Heliobakterid on roheliselt värvusega. Autotroofset kasvu pole neil ka õnnestunud näidata. Nende metabolism on fotoheterotroofne. Osad neist moodustavad endospoore ja see seletab nende hapniku- ja kuivataluvust. Dipikoliinhapet sisaldavad endospoorid on kirjeldatud Heliobacterium fasciculumil ja H. gestiil. See on ainus juhus, kus fotosünteesivatel anaeroobsetel bakteritel on kirjeldatud spoore. Nad fikseerivad ka N2. Märjas mullas (riisipöllud) vöivad heliobakterid koos tsüanobakteritega kindlasti taimi varustada lämmastikuga.
Perekonnad Pectinatus ja Megaspaera. Mölemad on villitud pastöörimata ölle saastajad. Esmalt kirjeldati neid 1970ndatel aastatel. Pöhjustavad ebameeldivat löhna ja maitset (H2S ja rasvhapped: butüraat ja propionaat). Pectinatus kirjeldati 1978. a. ja kuna ta ei sarnanenud teiste senikirjeldatutega, siis tehti ettepanek luua uus perekond Pectinatus. Liik oli P. cerevisiiphilus. Megasphaera isoleeriti esmalt Saksa öllest 1979. a. Pandi nimi M. cerevisiae. Praegu on neid kahte perekonda isoleeritud öllest igal pool maailmas. Kuna mölemad bakterid on anaeroobid ja öllevillimise tehnoloogia on sedavörd täiustunud, et üha vähem jäab öhku pudelitesse ja purki, siis on seal head tingimused nende bakterite arenguks, kui nad korra sinna on sattunud. Nad saastavad pastöriseerimata villitud õlut.
Köik senikirjeldatud Pectinatuse tüved on isoleeritud saastunud öllest. Veel on neid leitud pärmijuuretistest, villimisliinide pesuveest ja villimisliinide ja taarasäilitusruumide öhust. Kuigi need bakterid on anaeroobid, on nad vöimelised säiluma ka aeroobsetes tingimustes. Pectinatus Pectinatus cervisiiphilusel on raku ühel poolel viburid, mis meenutavad kammi. Rakud on veidi köverdunud, ümarate otstega, kas ühekaupa vöi paaris, harva ahelas. Elektronmikrofotodel meenutab Pectinatuse rakukest g(-) rakukesta, kuna välismembraan esineb. Tegelikult on tal aga väga paks peptidoglükaankiht. Selline rakukest nagu g(+) ja g(-) kesta vahepealne. Samatüüpi rakukest (paks peptidoglükaan + välismembraan on ka kiirgusresistentsel bakteril Deinococcus!).
Füsioloogia ja biokeemia Happelises ja kõrge alkoholisisaldusega (üle 5.2%) ölles ei kasva. Pectinatus ei suuda kasvada virde pöhilisel suhkrul-maltoosil, küll aga kasvablaktaadil. Seega stimuleerib piimhapebakterite esinemine ölles Pectinatust. Glükoosi kääritamisel moodustuvad propionaat, atsetaat, suktsinaat, laktaat ja atsetoiin. Köige rohkem tekib propionaati. Propionaat tekib samas rajas, nagu perekonnal Propionibacterium. Moodustab ka H2S, mis samuti rikub õlle maitset. Pectinatuse Isoleerimine. Kasutatakse söödet, mis sisaldab laktaati ja Pb-atsetaati. Pectinatus kasvab laktaadil ja toodab H2S, mis annab PbS ja mustad kolooniad. Inkubeeritakse ja külvatakse anaeroobsetes tingimustes.
Megasphaeraon g(-) värvuv kokk. Rakud on ühekaupa vöi lühikeste ahelatena. M. cerevisiae on õllesaastaja. Megasphaera isoleerimine. Megaspaera glükoosil ei kasva ja teda selekteeritakse välja fruktoosil vöi laktaadil. Füsioloogia ja biokeemia. Happelises ölles ja kanges ölles ei kasva. C-allikate ring on väga kitsas. Nad ei kasva glükoosil, küll aga kasvavad fruktoosil ja laktaadil. Käärimisproduktideka on butüraat, atsetaat, propionaat jne. Pöhiline saasteprodukt ölles on butüraat. Toodavad ka H2S. Rikuvad ära ölle maitse. Kuidas õlles tuvastada Megasphaerat ? Villitud pudeleid inkubeeritakse suletult 30oC juures 6 nädalat ja jälgitakse hägustumist. Vöib teha ka nii, et kohe pärast pudeli täitmist öllega lisatakse pudelisse kontsentreeritud söödet fruktoosiga, pudel suletakse ja inkubeeritakse 30oC juures 4 nädalat. Kui ölu hägustub, siis mikroskoobitakse ja nuusutatakse jne. Testida saab ka PCR-ga.
Perekond Selenomonas Need on anaeroobsed g(-) värvuvad, kuukujulised bakterid, kes liiguvad köhtmise viburite kimbu abil. Liikumine on tõmblev. Selenomonast kirjeldati juba möödunud sajandil, kus ta leiti suuöönest ja nimetati Spirillum sputigenumiks. Nüüd on ta nimi Selenomonas sputigena. Selenomonast on isoleeritud loomavatsast, inimese soolest ja suuöönest hambakatust. Vatsast isoleeritud tüved kuuluvad liiki Selenomonas ruminantium. See bakter toodab ka ureaasi ja saab vatsas lagundada uureat. Nad kääritavad lahustuvaid suhkruid ja laktaati propioonhappeks. Selenomonas ja igemehaigused Ilmselt vaatles esmalt neid Leeuwenhoek hambakaapest. Neid on inimese suus igemetaskutes ja tavaliselt on neid rohkem inimestel, kellel on igemehaigused. Selenomonase rakukesta lipopolüsahhariidid on toksilised koekultuuri rakkudele. Seega võivad nad olla paradontoosi põhjustajaks.
Perekond Sporomusa Perekond loodi 1984. a. Mölleri poolt. Ta kirjeldas graamneg., ovaalseid kuni banaanikujulisi pulkbaktereid, kes moodustasid endospoore. Spoorid on ümarad vöi ovaalsed, terminaalsed vöi subterminaalsed ja vöivad rakku paisutada. Rakukest on neil elektronmikroskoobis tüüpilise g(-) ehitusega ja sisaldab välismembraani. Värvub ka Grami järgi g(-), kuid peptidoglükaani struktuur on tal sarnane g(+) bakterite omale. Sporomusa ei hinga sulfaadiga ja ka see eristab teda Desulfotomaculumist. Pöhiline käärimisprodukt, mida ta moodustab, on atsetaat. Atsetaat tekib suhkrutest, org. hapetest ja alkoholidest, metanoolist, COst, CO2 + H2-st. Seega on ta atsetogeen. Leida vöib Sporomusat mageveekogude mudast, mullast, silost, loomasönnikust, termiitide soolestikust, kus ta toodab CO2 + H2-st atsetaati, mis on termiidile energiaallikaks.
Perekond Veillonella Siia kuuluvad väikesed (0.3-0.5 mkm) graamnegatiivselt värvuvad kokid. Vanas Bergey kästraamatud grupeeriti nad kokku anaeroobsete g(-) kokkidega. Rakukesta ehitustüüp ikkagi g(+). Rakud on tavaliselt paarides, liikumatud ja asporogeensed. Nad ei käärita suhkruid, sest neil puudub heksokinaas, küll aga kääritavad nad laktaati, püruvaati, malaati ja fumaraati. Pöhiline käärimisprodukt on propionaat ja atsetaat. Kus leidub? Loomade ja inimese seedetraktis, suuöönes, keelel, hambakatus, tupes. Kääritavad piimhapet, mida piimhapebakterid toodavad. Seetõttu on neid just seal, kus piimhapebaktereidki, näiteks suuõõnes.
Koloniseerimine Veillonella ise epiteelile ei kinnitu, kuid suudavad kinnituda teistele kinnitumisvöimelistele bakteritele, nagu Streptococcus, Fusobacterium, Actinomyces, Propionibacterium. Veillonella ilmub suuöönde varsti pärast lapse sündi. On näidatud, et Veillonella arvukus suus söltub Streptococcus salivariuse arvukusest. , kelle abil ta kinnitub ja kelle poolt toodetavat laktaati ta edasi kääritab. Streptokokkide rakuväline glükosüülitransferaas vöib seostuda V. parvula rakkudele ja sellised ensüümiga seotud veillonellad adsorbeeruvad juba hamba pinnale, kui seal esineb sahharoosi. Seda, et Veillonella vajab suuööne koloniseerimiseks Streptococcust, on näidatud ka gnotobiontsel roti mudelil. Kui rotte infitseeriti Veillonellaga, kolonisatsiooni ei toimunud, kui aga eelnevalt nakatati Streptococcusega ja siis Vellonellaga, siis Veillonella koloniseeris suuööne kiiresti.
Patogeensus. Inimesel vöivad pöhjustada ka pöletikke: hambajuure kanali pöletikke, sinusiiti, günekoloogilisi infektsioone, kopsupöletikku. Suuöönes on valdav V. parvula (varasem nimi Veillonella alcalescens). Seda bakterit on eriti palju igemepöletikuga inimestel. Igemerakkude koekultuuridel on näidatud, et propionaat on neile toksiline ja sellega saab seletada Veillonella poolt pöhjustatud igemekahjustusi.
Klostriidide klassi kuulub ka Epulopiscium Suur bakter on Epulopiscium fishelsonii, keda algselt peeti algloomaks. Tema pikkus on üle 600 µm ja ruumala miljon korda suurem, kui tavalistel bakteritel. Seega on ta suurem, kui näiteks kingloom. Ta elab troopilistes vetes ja on kalade (kirurgkala Acanthurus olivaceus) soolestiku sümbiont. Epulopiscium
Klostriidide klassi kuulub ka Metabacterium polyspora, kelle rakus on mitu endospoori. Fülogeneetiliselt on Metabacterium lähedane nii Epulopisciumiga kui ka klostriididega Metabacterium polyspora (vasakul) rakus võib moodustuda kuni 9 endospoori. Epulopiscium sp (paremal) rakus aga arvukalt tütarrakke. 16SrRNA järjestuste alusel on need kaks bakterit lähedased klostriididega
Fülogeneesipuul paiknevad Metabacterium ja Epulopiscium lähestikku klostriididega
Klass Mollicutes (mükoplasmad) Sinna kuulub 5 erinevat sugukonda. Olulisimad perekonnad on Mycoplasma, Ureaplasma, Spiroplasma, Anaeroplasma jmt. Selle rühma juures ka Erysipelothrix
Esmalt isoleeriti 1960. a. ühelt kopsupõletikuga haigelt ja hakati nimetama PPLO organismideks (pleuropneumonia like organisms). Traditsiooniliselt nimetatakse mükoplasmadeks baktereid, kes on väga väikesed ja kellel puudub rakukest, nad ei suuda sünteesida peptidoglükaani prekursoreid. Tänu kesta puudumisele on nad morfoloogiliselt pleomorfsed, st. ühes ja samas kultuuris vöib kohata ümaraid, pirnikujulisi, hargnenud ja helikaalseid rakke. Raku diameeter 0.3-0.8 mkm. Nad on väikseimad iseseisvalt paljunevad mikroorganismid. Mükoplasmad on pleomorfsed
Tänu rakukesta puudumisele on mükoplasmad osmootselt tundlikud, resistentsed penitsilliinile, tundlikud detergentidele ja tardsöötmel moodustavad härjasilmakolooniaid. Lihtsaim ehk väikseim senisekveneeritud genoom - kõigest 580 kb - kuulub Mycoplasma genitalium’ile (1998 a. seisuga). Temal on 482 geeni. Mükoplasma kolooniad
Mükoplasmad on kas saproobid, kommensaalid või parasiidid. Nad on kas fakultatiivsed anaeroobid või ranged anaeroobid. Vanas Bergey määrajas moodustasid mükoplasmad iseseisva klassi Mollicutes, mis oli ainus klass höimkonnas Tenericutes. Tenericutes oli üks neljast höimkonnast riigis Prokaryotae. Praegu arvatakse, et ilmselt on mükoplasmad arenenud degeneratiivse evolutsiooni teel g(+) organismidest, kel on madal GC% DNA-s.
Mükoplasmad on enamasti liikumatud ja vibureid ei oma, osa siiski suudab tardsöötmel libiseda, nende hulgas ka inimese ja loomade patogeene. Libisevatel mükoplasmadel on rakkudel leitud spetsiaalne tipustruktuur, nagu iminapake, mille abil ta saab kinnituda peremeesrakule ja roomamine toimub alati, napp ees. Mycoplasma genitalium. Temal on ka senisekveneeritud bakteritest väikseim genoom
Kus esinevad? Inimese, loomade, putukate ja taimede parasiidid. Inimese ja loomade puhul paiknevad nad hingamisteede ja urogenitaaltrakti limaskestadel, ka silma, söögitoru epiteelil, rinnanäärme juhade epiteelil ja mönede loomade liigestes. Obligaatsed anaeroobid Anaeroplasma ja Asteroplasma on seni leitud vaid lamba ja veise vatsast, kus nad kääritavad tärklist. Spiroplasmasid on putukate süljenäärmetes ja sooles. Sealt nad satuvad taimedesse, kus nad pöhjustavad haigusi.
Molekulaarbioloogia ja geneetika Väike genoom, järelikult vähe geene. Erinevaid geene on mükoplasmadel ca 5x vähem, kui E. coli’l. Mükoplasmad on väikseimate genoomidega bakterid. DNA GC- sisaldus on madal: 23 - 41 %. Kuna genoom on väike, siis sünteesitakse ka vähem valkusid ja töesti on mükoplasmad metaboolselt defektsed, mis on seotud nende parasiitse eluviisiga. Nende keeruliste toitumisnöudluste töttu on neid laboratoorsetel söötmetel seetöttu raske kasvatada. Mükoplasmadel puuduvad tsütokroomid, tsitraaditsükli ensüümid ning enamik biosünteesiradadest. Näiteks M. genitaliumi ja Haemophilus influenzae genoomide võrdlemine näitas, et kui H. influenzae’lon aminohapete biosünteesi tarvis 68 geeni, M. genitalium’il ainult üks. Kofaktorite biosünteesiks on M. genitalium’il 5 geeni H. influenzae 54 vastu.
Mõned mükoplasmad vajavad kasvuks kolesterooli(ebatavaline bakterite puhul), mida nad lülitavad membraanidesse. Selline tsütoplasmamembraan on tugevam, kui ilma steroolideta membraan. See on ka arusaadav, sest mükoplasmadel kesta pole. Steroolidevajaduse järgi jagataksegi mükoplasmad 2 gruppi: steroole vajavad ja mittevajavad perekonnad. Steroole ei vaja Acholeplasma ja Asteroplasma. Neil mükoplasmadel, kel steroole membraani ei lülitata, on membraanis karotinoidid, mis tugevdavad seda. Osadel mükoplasmadel on membraanis lipoglükaanid, mis meenutavad ehituselt lipopolüsahhariide, kuid lipiid A puudub neis. Ka see teeb membraanid tugevamaks ja võib aidata ka seostuda mükoplasmadel eukarüootsetele rakkudele (rakkude pinnaretseptoritele). Nii nagu lipopolüsahariididki on lipoglükaanid antigeensed ja nende vastu moodustuvad antikehad. Mõned mükoplasmad vajavad ka uureat. Osad kasutavad biokeemiliste reaktsioonide energiaallikana ATP asemel pürofosfaati.
Mükoplasmade genoome iseloomustab väga madal GC-sisaldus (23-41%). Madalaim 23 %-line GC-sisaldus on teoreetiliselt minimaalne väärtus, mis veel võimaldab kodeerida normaalse aminohappesisaldusega valke. Koodoni 3. positsioonis kasutavad mükoplasmad enamasti A-d või U-d. Mycoplasma ja Spiroplasma liigidkasutavad universaalset stop-koodonit UGA trüptofaani koodonina nagu mitokondergi. Acholeplasma’son UGA stop-koodoniks, trüptofaani kodeerib UGG nagu tavaliselt. Selle erineva koodonikasutuse töttu ei saa mükoplasmadest kloneeritud geene ekspresseerida E. coli’s. E. coli ei lülita UGA kohal valku Trp, vaid termineerib ahela sünteesi. Tekivad köndistatud valgud.
Füsioloogia Mükoplasmad jagatakse fermentatiivseteks ja mittefermentatiivseteks. Mittefermentatiivsetel mükoplasmadel puudub heksokinaas, fosfofruktokinaas ja aldolaas ja EMP rada neil ei funksi. Küll aga saavad nad kasutada rasvhappeid ja alkohole ja neid aeroobselt lagundada. Köigil seniuuritud mükoplasmadel on väga puudulik hingamissüsteem, neil puuduvad kinoonid ja tsütokroomid, seega ei suuda nad oksüdatiivselt fosforüülida. Fermentatiivsetel mükoplasmadel moodustub ATP glükolüüsis substraatsel fosforüülimisel. Mittefermentatiivsetel võib ATP moodustuda arginiini dihüdrolaasi reaktsioonis. Neil on membraanis aktiivne karboksüpeptidaas, mis vöib peremeesraku valkudest vabastada Arg ja varustada mükoplasmat energiaallikaga. L-Arg → L-Tsitrulliin → L-Orn + karbaomüül-P → ATP
Uureat lagundavatel mükoplasmadel pole leitud Arg dihüdrolaasi, glükolüüsi ega atsetaadi kinaasi. Seega peab neil olema mingi muu vöimalus ATP tootmiseks. On esitatud hüpotees, mille järgi uurea hüdrolüüsil moodustuv NH3 suunatakse rakust välja ja moodustub ioongradient ja membraanpotentsiaal, mille energia arvel sünteesitakse ATPd.
Patogeensus M. pneumoniae pöhjustab farüngiiti, trahheobronhiiti, kopsupöletikku, artriiti ja konjunktiviiti. Kopsupõletikku põhjustavad just koolilastel ja noorukitel. Ureaplasma urealyticum pöhjustab meestel mittegonokokilist uretriiti, neerukivide teket ja naistel loote emakasisesid infektsioone. M. hominisvõib põhjustada püelonefriiti ja väikese vaagna põletikke (pelvic inflammatory disease). Seotud viljatusega. Mükoplasmad on tülikad koekultuuride saastajad. Seerum, millel kultuure kasvatatakse, vöib olla saastunud, kuid saastus vöib olla pärit ka koekultuuriga töötaja suust (süljepiisad).
Patogeneesi mehhanismid Inimest ja loomi nakatavad mükoplasmad kleepuvad tugevasti limaskestadele hingamiselundites ja urogenitaaltraktis ja enamasti ei tungi verre ega kudedesse. Seega nad on pinnaparasiidid. Adhesioon on piisavalt tugev, et vältida mükoplasmade väljauhtumist. Mükoplasmal peab adhesiiniks olema möni rakumembraani komponent (näiteks lipoglükaan). Tema membraan kleepub peremeesraku membraanile. See tihe kontakt toob kaasa selle, et mükoplasma poolt toodetud NH3, H2O2 ja superoksiidradikaalid saavad kergesti tungida peremeesrakku ja kahjustada selle kudesid. Mükoplasmad epiteeli pinnal
Mükoplasmade poolt toodetud H2O2 ja superoksiidradikaal tungivad peremeesrakku. Superoksiidradikaal inhibeerib peremeesraku katalaasi, mis viib H2O2 kuhjumisele ja see omakorda pöhjustab SODi tagasisidestusliku inhibitsiooni. See toob kaasa rakus H2O2 ja superoksiidradikaali kontsentratsiooni töusu ja membraanid saavad oksüdatiivselt kahjustatud.
Perekond Spiroplasma Varem peeti spiroplasmasid kas viirusteks vöi spiroheetideks. See on ka arusaadav, sest nad läbivad bakterifiltreid. Perekonna tüüpliik on Spiroplasma citri. Spiroplasmad on liikuvad, helikaalsed, kestata. Kultuuris on alati ka kokoidseid rakke. Spiroplasmadel on leitud rakusiseseid fibrille, mis arvatakse osalevat liikumises. Spiroplasmad on tüüpilised putukate ja puukide parasiidid, mis esinevad sooles, vahel ka hemolümfis ja süljenäärmetes. Spiroplasmad vöivad putukale ka surmavad olla, juhul kui nad soolest tungivad hemolümfi ja pöhjustavad septitseemiat. Selliseid juhte on kirjeldatud mesilastel, kes on nakatatud S. apise vöi S. melliferumi poolt. Spiroplasmadest pöhjustatud taimehaigusi esineb apelsinipuudel, maisil, sidrunipuudel jpt taimedel: kapsas, redis, lillkapsas, salat, kirss, pirn. Spiroplasmad paiknevad taimede söeltorudes ja taim kärbub.
Perekond Erysipelothrix Tuntuim liik E. rhusiopathiae. Varem klassifitseeriti neid korünebakterite hulka. Väike sale g(+) pleomorfne pulkbakter. Liikumatu. Mikroaerofiil. Põhjustab punataudi. Siseneb inimese organismi kriimustuse kaudu. Satub sinna peamiselt kalast või lihast, aga ka pinnasest. Punataud esineb sigadel, lammastel, veistel, küülikutel. Paljud loomad ja linnud kannavad seda bakterit soolestikus. Inimesel on punataud mitme vormina, põhiline on põletikuline nahakahjustus. Nahk punetab, sügeleb, valutab. AB ravi (penitsilliin) aitab. On ka raskemaid punataudi vorme (sepsis näiteks).
Klass Bacilli (batsillid) Siia kuulub kaks seltsi: Bacillales ja Lactobacillales Selts Bacillales Suurim perekond on Bacillus (114 liiki), kuigi osa liike on perekonna Bacillus alt viidud uutesse perekondadesse. On tehtud juurde näiteks perekonnad Paenibacillus, kuhu on kuuluvad näiteks P. polymyxa ja P. macerans. Nad mõlemad fikseerivad N2. Perekonda Bacillus kuuluvad aeroobsed ja ka mõned fakult. anaeroobsed sporogeensed pulkbakterid. Batsillid toodavad ka antibiootikume: batsitratsiini, polümüksiini, gramitsidiini, tsirkuliini.
1835. a. kirjeldas Ehrenberg bakteri Vibrio subtilis, mis nimetati 1872. a. ümber Bacillus subtiliseks Cohni poolt. Bacilluse olulisim omadus on endospooride moodustamine. Esmalt kirjeldatigi spoore B. subtilisel (Ferdinand Cohn), seejärel aga B. anthracisel (Robert Koch). Koch kirjeldas B. anthracisel ka spoor-vegetatiivne rakk-spoor tsüklit. Sporangium spooriga on hästi näha faaskontrastmikroskoobis, sest ta murrab tugevast valgust. Ka Grami järgi värvimisest piisab, et spoori rakus näha: ta jääb värvusetuks. Kui mulda, vett, muda vöi ka toiduaineid kuumutada 80oC juures 10-30 min, et tappa veget. rakke, tsüste ja eksospoore ja seejärel välja külvata sobivale söötmele, siis on kindlasti vöimalik isoleerida endospoore moodustavaid baktereid. Selektiivsel isoleerimisel saab kombineerida erinevaid C-allikaid, söötme pHd, kasvutemperatuuri jne, et isoleerida erinevaid sporogeene. B. subtilise endospoorid
Selts Lactobacillales Selts Lactobacillales Selts Bacillales
Perekond Bacillus Perekond Bacillus on füsioloogiliselt ja geneetiliselt suhteliselt kirju. Perekonnas on siiski ka homogeenseid klastreid : Näiteks liigid B. anthracis, B. cereus ja B. thuringensis on kõik lähedased ja omavahel 80-100% DNA hübridiseerimise järgi homoloogsed. Seetõttu on neid soovitatud kõik kanda liigi B. anthracis alla alamliikidena. Nende kolme liigiga on sarnane ka B. mycoides. Avirulentne B. anthracis on v. sarnane B. cereusega. Batsillide seas on ka teisi homoloogseid klastreid. B. amyloliquefaciens ja B. subtilis on 51-81 homoloogsed. Nendega on sarnane ka B. natto. Biotehnoloogiliselt on oluline B. subtilis kui amülaasi tootja. Meditsiiniliselt on olulised B. anthracis ja B. cereus.
B. anthracis ja siberi katk e. antraks. 1876. a. Koch vaatles B. anthracist siberi katku surnud loomade veres ja nakatas ka terveid loomi (hiiri) batsillide puhaskultuuridega ja näitas ära, et ilmnesid siberi katku tüüpilised tunnused. Pasteur töötas välja vaktsiini siberi katku vastu ja korraldas ka selle demonstreerimiseks avaliku katse. Vaktsiin oli 43oC juures kasvatamisega nõrgestatud tüvi. B. anthracis moodustab aheldunud pulkadest niite, millel on paks kapsel. Raku otsad tömbid. Niidid meenutavad mikroskoobi all bambuskeppe. Spoorid säiluvad väga hästi, eriti külmas. Keetmine hävitab nad ca 10 minutiga.
B. anthracis ja siberi katk e. antraks. 1979.a. oli Siberis Sverdlovski lähedal suur siberi katku epideemia, kus haigestus üle 1000 inimese. Arvatakse, et see oli seotud mingi önnetusega bakterrelva tootvas söjatehases. Bacillus anthracis on hea bakterioloogiline relv, sest ta põhjustab haigestumist nii inimesel kui ka tema koduloomadel. Teda on odav kasvatada, ta sporuleerub kergesti, spoorid püsivad kaua nakatamisvõimelisena, neid on võimalik rakettidega kohale toimetada. Iraak on tootnud üle 2000 galloni siberi katku tekitaja kultuuri, valmistanud sellest 50 bakterpommi.
B. anthracise spoorid on väga termoresistentsed, taluvad hästi ka kuivust, desinfitseerivaid aineid jne. Seetöttu vöivad nad säiluda mullas aastakümneid. Ka vihmaussid ja raipesööjad linnud vöivad spoore levitada. Primaarselt on siberi katk rohusööjate loomade haigus. Loomad nakatuvad, kui spoorid satuvad nende verre näiteks vigastuste kaudu suuööne epiteelis või ka hingamisteede kaudu. Haiguse ägedas vormis ilmnevad verejooksud, kopsuturse jms.
Ka inimesed nakatuvad siberi katku Neil vöib esineda haiguse nahavorm (pildil), mis tekib siis, kui inimene töötleb haigete loomade nahku, keresid jne. Inimestel on levinuim just nahavorm. Spoorid satuvad nahavigastuse kaudu epidermise alumisse kihti, idanevad ja toodavad toksiini. Tekib punetus, sellest mädavill, mis haavandub ja kattub musta koorikuga. Villi ümber tekib tugev turse. Ravi puudumisel läheb haigus üle sepsiseks. Kui ei ravita, on suremus 20%. Valikravimiks penitsilliin. Kui aga süüakse haigete loomade halvasti küpsetatud liha, siis areneb välja haiguse soolevorm. Seda esineb inimestel v. harva.