Download
1 / 1
20 likes | 159 Views
Structure and Properties of Blue Copper Azurin Under Applied Compression : Simulation Studies Andrew Hung 1 , Jian-wei Zhao 2 , Jason J. Davis 2 and Mark S.P. Sansom 1 1 Department of Biochemistry, University of Oxford, OX1 3QU, United Kingdom

E N D
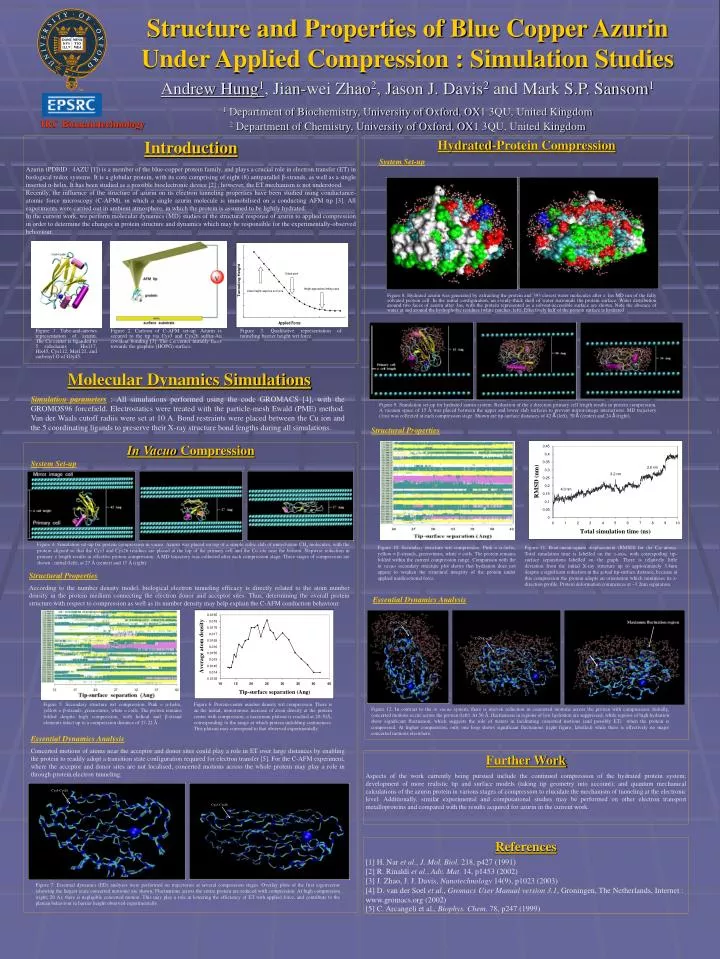
Structure and Properties of Blue Copper Azurin Under Applied Compression : Simulation Studies Andrew Hung1, Jian-wei Zhao2, Jason J. Davis2 and Mark S.P. Sansom1 1 Department of Biochemistry, University of Oxford, OX1 3QU, United Kingdom 2 Department of Chemistry, University of Oxford, OX1 3QU, United Kingdom IRC Bionanotechnology Introduction Azurin (PDBID : 4AZU [1]) is a member of the blue-copper protein family, and plays a crucial role in electron transfer (ET) in biological redox systems. It is a globular protein, with its core comprising of eight (8) antiparallel β-strands, as well as a single inserted α-helix. It has been studied as a possible bioelectronic device [2] ; however, the ET mechanism is not understood. Recently, the influence of the structure of azurin on its electron tunneling properties have been studied using conductance-atomic force microscopy (C-AFM), in which a single azurin molecule is immobilised on a conducting AFM tip [3]. All experiments were carried out in ambient atmosphere, in which the protein is assumed to be lightly hydrated. In the current work, we perform molecular dynamics (MD) studies of the structural response of azurin to applied compression in order to determine the changes in protein structure and dynamics which may be responsible for the experimentally-observed behaviour. Hydrated-Protein Compression System Set-up Figure 8. Hydrated azurin was generated by extracting the protein and 393 closest water molecules after a 1ns MD run of the fully solvated protein cell. In the initial configuration, an evenly-thick shell of water surrounds the protein surface. Water distribution around two faces of azurin after 3ns, with the protein represented as a solvent-accessible surface are shown. Note the absence of water at and around the hydrophobic residues (white patches; left). Effectively half of the protein surface is hydrated Figure 1. Tube-and-arrows representation of azurin. The Cu center is liganded to 5 sidechains : His117, His45, Cys112, Met121, and carbonyl O of Gly45. Figure 2. Cartoon of C-AFM set-up. Azurin is secured to the tip via Cys3 and Cys26 sulfur-Au covalent bonding [3]. The Cu center initially faces towards the graphite (HOPG) surface. Figure 3. Qualitative representation of tunneling barrier height wrt force Molecular Dynamics Simulations Simulation parameters : All simulations performed using the code GROMACS [4], with the GROMOS96 forcefield. Electrostatics were treated with the particle-mesh Ewald (PME) method. Van der Waals cutoff radiis were set at 10 A. Bond restraints were placed between the Cu ion and the 5 coordinating ligands to preserve their X-ray structure bond lengths during all simulations. Figure 9. Simulation set-up for hydrated azurin system. Reduction of the z direction primary cell length results in protein compression. A vacuum space of 15 Å was placed between the upper and lower slab surfaces to prevent mirror-image interactions. MD trajectory (1ns) was collected at each compression stage. Shown are tip-surface distances of 42 Å (left), 30 Å (center) and 24 Å (right). Structural Properties In Vacuo Compression System Set-up Figure 4. Simulation set-up for protein compression in vacuo. Azurin was placed on top of a simple cubic slab of united-atom CH4 molecules, with the protein aligned so that the Cys3 and Cys26 residues are placed at the top of the primary cell and the Cu site near the bottom. Stepwise reduction in primary z length results in effective protein compression. A MD trajectory was collected after each compression stage. Three stages of compression are shown : initial (left), at 27 Å (center) and 17 Å (right). Figure 10. Secondary structure wrt compression. Pink = -helix, yellow = β-strands, green=turns, white = coils. The protein remains folded within the current compression range. Comparison with the in vacuo secondary structure plot shows that hydration does not appear to weaken the structural integrity of the protein under applied unidirectional force. Figure 11. Root-mean-square displacement (RMSD) for the C atoms. Total simulation time is labelled on the x-axis, with correspoding tip-surface separations labelled on the graph. There is relatively little deviation from the initial X-ray structure up to approximately 3.4nm despite a significant reduction in the actual tip-surface distance, because at this compression the protein adopts an orientation which minimises its z-direction profile. Protein deformation commences at ~3.2nm separation. Structural Properties According to the number density model, biological electron tunneling efficacy is directly related to the atom number density in the protein medium connecting the electron donor and acceptor sites. Thus, determining the overall protein structure with respect to compression as well as its number density may help explain the C-AFM conduction behaviour. Essential Dynamics Analysis Cys3-Cys26 Maximum fluctuation region Cys3-Cys26 Cys3-Cys26 Figure 5. Secondary structure wrt compression. Pink = -helix, yellow = β-strands, green=turns, white = coils. The protein remains folded despite high compression, with helical and -strand elements intact up to a compression distance of 27-22 Å Figure 6. Protein-center number density wrt compression. There is an the initial, monotonous increase of atom density at the protein center with compression; a maximum plateau is reached at 20-30Å, corresponding to the range at which protein unfolding commences. This plateau may correspond to that observed experimentally. Figure 12. In contrast to the in vacuo system, there is uneven reduction in concerted motions across the protein with compression. Initially, concerted motions occur across the protein (left). At 30 Å, fluctuations in regions of low hydration are suppressed, while regions of high hydration show significant fluctuation, which suggests the role of waters in facilitating concerted motions (and possibly ET) when the protein is compressed. At higher compression, only one loop shows significant fluctuaions (right figure; labelled) while there is effectively no major concerted motions elsewhere. Essential Dynamics Analysis Concerted motions of atoms near the acceptor and donor sites could play a role in ET over large distances by enabling the protein to readily adopt a transition state configuration required for electron transfer [5]. For the C-AFM experiment, where the acceptor and donor sites are not localised, concerted motions across the whole protein may play a role in through-protein electron tunneling. Further Work Aspects of the work currently being pursued include the continued compression of the hydrated protein system; development of more realistic tip and surface models (taking tip geometry into account); and quantum mechanical calculations of the azurin protein in various stages of compression to elucidate the mechanism of tunneling at the electronic level. Additionally, similar experimental and computational studies may be performed on other electron transport metalloproteins and compared with the results acquired for azurin in the current work. Cys3-Cys26 Cys3-Cys26 References [1] H. Nar et al., J. Mol. Biol. 218, p427 (1991) [2] R. Rinaldi et al., Adv. Mat. 14, p1453 (2002) [3] J. Zhao, J. J. Davis, Nanotechnology 14(9), p1023 (2003) [4] D. van der Soel et al., Gromacs User Manual version 3.1, Groningen, The Netherlands, Internet : www.gromacs.org (2002) [5] C. Arcangeli et al., Biophys. Chem. 78, p247 (1999) Figure 7. Essential dynamics (ED) analyses were performed on trajectories at several compression stages. Overlay plots of the first eigenvector (showing the largest scale concerted motions) are shown. Fluctuations across the entire protein are reduced with compression. At high compression (right; 20 A), there is negligible concerted motion. This may play a role in lowering the efficiency of ET with applied force, and contribute to the plateau behaviour in barrier height observed experimentally.






















