Download
1 / 17
190 likes | 313 Views
From Rost et al., “Plant Biology”, 2 nd edn. Photosynthesis: the light-independent reactions. Biol 3470 Plant Physiol Biotech 5.5 to 5.12 Lecture 9 Thurs. Feb. 2, 2006. The process of carbon fixation in plants goes by many names. Including: The dark reactions

E N D
From Rost et al., “Plant Biology”, 2nd edn Photosynthesis: the light-independent reactions Biol 3470 Plant Physiol Biotech 5.5 to 5.12 Lecture 9 Thurs. Feb. 2, 2006
The process of carbon fixation in plants goes by many names • Including: • The dark reactions • The enzymatic reactions of photosynthesis • Reductive pentose phosphate cycle • C3 cycle • The photosynthetic carbon reduction (PCR) cycle (in the textbook)
The PCR cycle converts atmospheric carbon to organic molecules • Convert CO2 to stable phosphorylated carbon intermediates (specifically- a three-carbon carbohydrate, 3-PGA) • Uses the energy produced in the light-dependent reactions to reduce CO2 • Convert less complex → more complex molecules • Fight entropy • Pathway elicited in late 1940s and early 50s by U.S. plant physiologist Melvin Calvin using labeled 14CO2 • feed plants 14CO2 • Allow metabolism • Kill, extract, examine small carbohydrates that contain 14C using paper chromatography and autoradiography
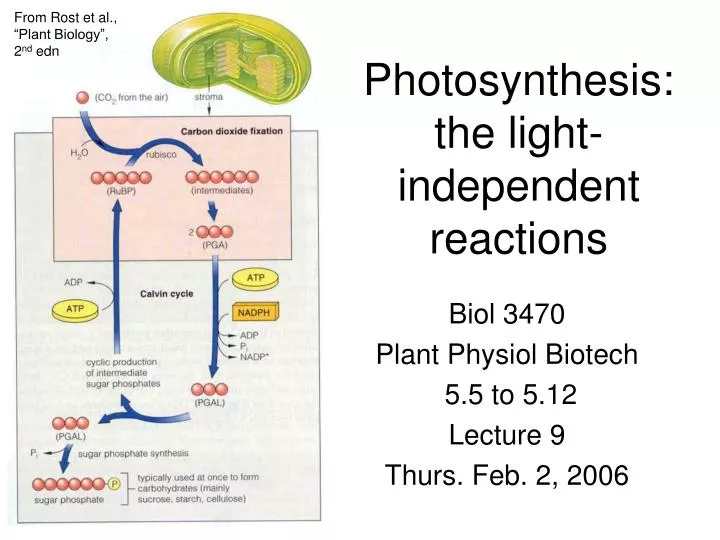
The PCR cycle contains 3 distinct segments Fig. 5.8 Step 1: Carboxylation fixes CO2 using the enzyme rubisco • 14CO2 fixed first into 3-phosphoglycerate (3 Cs ≡ C3 cycle) • 3-PGA is the first organic product of the PCR cycle • Given this product and reactant, one would assume the plant substrate of the PCR cycle would have ___Cs 1 3 2 Rubisco reaction • Actually, the plant substrate for the PCR cycle is a five-carbon substrate, RuBP Unstable intermediate is hydrolyzed (2x ) Fig. 5.9
The key enzyme regulating carbon uptake by the PCR cycle is rubisco Rubisco • Enzyme with a high affinity for CO2 • Present in high amounts in the chloroplast stroma • Its activity maintains a CO2 gradient from the atmosphere • ΔGº′ = -35 kJ/mol (energetically favourable to occur spontaneously) • But its activity requires ATP + NADPH made in the light reactions elsewhere in PCR cycle • e.g. in 2nd step: reduction of 3-PGA to G3P • This is Step 2: Reduction Phosphorylate! Reduce! Fig. 5.10
The final step in the PCR cycle regenerates the rubisco substrate • This is accomplished via Step 3: Regeneration • Requires 1 ATP per CO2 • Note that the PCR cycle is autocatalytic • This means that it operates more quickly if CO2 and/or RuBP pools are low (e.g. in the morning, when the RuBP supply is depleted)
PCR cycle activity must be integrated with plant carbon metabolism as a whole • These include respiration (glycolysis) and macromolecule synthesis (for nucleic acids, lipids, carbohydrates, proteins) • Thus, the PCR cycle activity must be regulated by a number of mechanisms • The plant wants to keep CO2 fixation rate high to make more organic carbon • The carbon from 5 of every 6 molecules of G3P needs to be recycled to make RuBP and keep the cycle spinning • Only around one-sixth of the carbon fixed is exported from the leaf and supports growth and metabolism • To keep high CO2 fixation, the plant can prevent G3P export 2. Reduction Rubisco 1. Carbox-ylation Autocatalytic RuBP regeneration 3. Regener-ation The PCR cycle consumes the ATP and NADPH produced in the light-dependent reactions Fig. 5.11
Mechanisms of regulation of PCR cycle activity • The activity of rubisco is regulated by light • Complex mechanism driven by uptake of protons by thylakoid lumen between • Mg2+→ moves lumen → stroma to compensate for H+ uptake by thylakoids in light inactive active • Stromal pH ↑ activates rubisco • CO2→ binds to activating site on rubisco (not active site!) ≡ CARBAMYLATION • pH increase favours carbamylation (H+ sink in lumen) (light) Dark stroma pH = 5.0 Light Rubisco is now catalytically ready to fix atmospheric CO2! Fig. 5.14
Plant cells also respire: convert O2→CO2 (mt) • Via mitochondiral respiration at night • This is oxidative phosphorylation to generate ATP in the dark (R on diagram) • This also happens in the light! • Via rubisco → can use O2 as a substrate in photorespiration (PR) • Therefore, measuring NET gas exchange in photosynthetic organs is difficult! • We can define an apparent photosynthesis rate = CO2 fixation rate – CO2 evolution rate =gross p’syn – (mt R + PR) • At a low atmospheric [CO2] these values (GP) and (R+PR) are equal • this is the CO2 compensation point (Rubisco CO2-ase) (mt) (mt) (Rubisco O2-ase) Fig. 5.16
Photorespiration is due to rubisco’s oxygenase activity Fig. 5.18 • Makes 2-phospho-glycolate (2C) + 3-PGA from RuBP + O2 • This C in 2-P-glycolate not wasted but reassimilated by exchange of intermediates with 2 other organelles • Peroxisome • Mitochondria Exported or recycled to regenerate RuBP
The function of photorespiration is not immediately obvious Energetically wasteful, so why do it? Thoughts and theories… • [O2] in atmosphere has been low during most of evolutionary history • Therefore PR is an evolutionary relic? • No! PR mutants are lethal! → Therefore, PR is essential • No evolutionary pressure to get rid of O2-ase function • The salvage cycle does a good job of recovering photorespired C • Each 2 turns of the 2-P-glycolate salvage cycle forms 4 3-PGA • 1 lost, 3 returned to the PCR cycle • Complex salvage pathway works well! • Metabolic safety valve? • PR protects against photoxidative damage by allowing P.E.T. to continue at low [CO2] • e.g., under high light + low CO2 (photoinhibitory conditions, stomata closed, water-stressed)
The chloroplast oxidative pentose phosphate cycle allows plants to make NADPH in the dark Fig. 5.20 • Shares intermediates with PCR • Both at once: FUTILE CYCLE! • Use 3 ATP • No CO2 fixation! • Both pathways are light-regulated PCR a/k/a RPPC OPPC • Light induces changes the structure of the disulfide bonds of the pathways’ enzymes • PCR cycle enzymes active when reduced • OPPC cycle enzymes active when oxidized • Why have an OPPC? • Make NADPH in dark • Make ribose and deoxyribose for nucleic acid synthesis
How can plants minimize PR and maximize GP? Plants are separated into 2 main groups based on their ability to do this: • Plants where 3-C 3-PGA is product of CO2 fix’n = C3 • Plants where 4-C oxaloacetate is product of CO2 fix’n = C4 • C4 plants have 2 distinct photosynthetic tissues • Leaf anatomy differs from C3 leaf • Mesophyll • fewer chloroplasts • Bundle sheath cells • lots of chloroplasts • surround vascular tissue • thick cell walls prevent diffusion of CO2 out of BS cells and traps photorespired CO2 • No mesophyll cells are more than 2-3 cells away from BS • This ensures quick export of fixed CO2 as sucrose • Many chloroplasts needed to fix high [CO2] Fig. 5.21: C4 leaf X-section (e.g., maize)
C4 plants are present in all 18 plant families • This includes flowering plants as well C4 plants are: • Better at CO2 fixation (up to 3X more efficient) • Better at drought stress • Concentrate CO2 at rubisco active site and thus minimize CO2 loss! • How do they do this? • Fix CO2 into C4 organic acid in the mesophyll cell using PEP carboxylase (not rubisco!) • Use a transporter to move the acid into the bundle sheath cell • Release CO2 there • Fix CO2 via PCR • Recycle C3 acid released (pyruvate) back to mesophyll PEP carboxylase malate malate Malic Enzyme pyruvate pyruvate Fig. 5.22: the C4 carbon fixation pathway
Using the C4 pathway to fix carbon is not always an advantage for the plant • C4 metabolism pluses: • CO2 outcompetes O2 at rubisco active site: Less PR! • Much lower compensation point • maintain high CO2 fixation rates when stomata are partially closed → conserves H2O • Lower transpiration ratio = less H2O transported per CO2 assimilated • C4 metabolism minuses: • Need to “spend” 2 ATP per CO2 to recycle C3 acid back to the mesophyll cells • C3 plants often have an ecological advantage • Grow better in cooler climates and low irradiance • Higher CO2 assimilation rate in environments with lots of water
How do plants grow in the desert? • Use CAM metabolism: conserves H2O • CAM plants have an inverted stomatal cycle • Night: open • Day: closed • Therefore CO2 uptake at night→ accumulate malate in vacuole • During day → convert malate to starch via PCR cycle • Need PEPC as in C4 photosynthesis • Requires lots of PEP (PEPC substrate), provided from glycolytic breakdown of starch • CAM is similar to C4, but: • No specialized anatomy (specialized cell types) • No closed cycle of carbon intermediates Night Day PEPC Malic enzyme Decarbox- ylation export Large, watery Fig. 5.26
CAM plants are evolutionarily adapted to live in low water environments • CAM plants have even lower transpiration ratios than C4 plants BUT • Only fix <1/2 C of C3 and <1/3 of C4 plants → slow growers • But can • continue CO2 uptake under H2O stress • Reassimilate respired CO2 • Some plants can “switch on” CAM metabolism (facultative vs. obligatory) From http://www.arizonensis.org/images/plantae/cereus_gigant.jpg






















