Download

1 / 25
250 likes | 314 Views
Homeostatic plasticity and postsynaptic receptor trafficking.

E N D
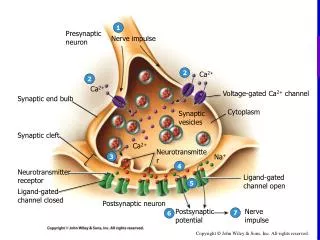
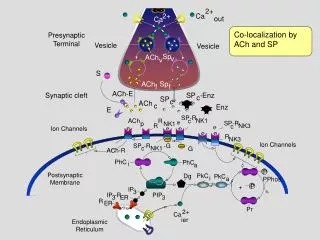
Homeostatic plasticity and postsynaptic receptor trafficking • Alterations in synaptic strength can be encoded presynaptically as alterations in the machinery releasing the neurotransmitter glutamate, or postsynaptically by changing the number or function of receptors sensing the glutamate signal. • A key advance has been the discovery that glutamate receptors are dynamically transported to and from the postsynaptic membrane. Such dynamic transport operates over a wide range of timescales and responds to diverse stimuli, matching the varied spectrum of plasticity modes. • It is now widely accepted that the trafficking of AMPA-type glutamate receptors mediates rapid synaptic modification, long term potentiation (LTP) and long term depression (LTD). • Synaptic scaling is a homeostatic form of plasticity that tends to restore neurotransmission activity to ‘baseline’ levels. • NMDA receptor scaling, as for that of AMPA receptors, occurs through changes in the number of receptors clustered at the synapse.
Disruption of AMPA receptor GluR2 clusters following long-term depression induction in cerebellar Purkinje neuronsShinji Matsuda, Thomas Launey, Sumiko Mikawa, and Hirokazu Hirai
Fig. 1. Induction of cerebellar LTD by PKC activation. • Whole-cell voltage-clamp recording was performed from a Purkinje cell at 25 days in vitro. Excitatory PostSynaptic Current(EPSC) was evoked by extracellular stimulation of a single granule cell. (A) Amplitudes of the evoked EPSC, plotted as a function of time relative to the beginning of TPA application (200 nM TPA at 0 min). Each data point represents the amplitude of one evoked EPSC, as a percentage of the control amplitude. The average of the EPSC amplitude before TPA application was set at 100%. Insets show the representative current traces taken at times (a), (b) and (c), respectively. (B) Each point represents the normalized EPSC amplitude from five Purkinje cells (recording condition as above). The EPSC amplitudes were averaged over a 2 min time window.
Using heterologous expression, the authors have shown previously that PKC phosphorylates Ser880 in the C-terminal domain of GluR2, and that this phosphorylation reduces the affinity of the receptor to the GRIP fragment containing PDZ 4–5 (amino acids 419–673) (GRIP419–673) (Matsuda et al., 1999), a region essential and sufficient for binding to GluR2 (Dong et al., 1997). • These results suggest that phosphorylation of Ser880 in GluR2 may regulate the postsynaptic localization of AMPA receptors by inhibiting GRIP binding.
Fig. 2. Effect of PKC activation on the co-localization of GluR2 and GRIP in transfected HEK 293 cells. • (A and B) HEK cells expressing either GluR2 (FITC labeled) (A) or GRIP419–673 (rhodamine labeled) (B). (C and D) HEK cells simultaneously expressing GluR2 and the GRIP fragment prior to (C) and after (D) TPA stimulation (200 nM, 20 min). (E) A cell expressing GluR2 and the GRIP419–673 20 min after treatment with TPA and GF109203X (200 nM), a selective PKC inhibitor. (F) A cell expressing mutant GluR2 (S880D) and GRIP419–673. • These results indicate that phosphorylation of Ser880 regulates the subcellular localization of GluR2 by controlling its binding to GRIP.
Fig. 3. Disruption of GluR2 clusters in response to PKC activation. (A) Co-localization of GluR2 and a presynaptic marker synaptophysin. Arrows indicate the spots representing immunoreactivity for both antibodies. Arrowheads indicate the GluR2- but not synaptophysin-immunoreactive spots. (B and C) Disruption of the postsynaptic GluR2 clusters following TPA application. Purkinje cells were treated with 200 nM TPA for 20 min, followed by staining with anti-GluR2 N-terminal antibodies (B) or rhodamine–phalloidin (C). (D) Phosphorylation of Ser880 in GluR2 following application of TPA. The membrane fraction of cultured Purkinje cells treated or not treated with TPA (200 nM, 20 min) was subjected to SDS–PAGE and immunoblotting with anti-GluR2 N-terminal antibodies (a) or antibodies specific for the phosphoSer880 residue in GluR2 (anti-P-Ser880 antibodies) (b). (E) Parallel decrease in the affinity of GluR2 for GRIP. Immunoprecipitation was performed using polyclonal anti-GRIP antibodies. Immunoprecipitated GRIP (a) and co-immunoprecipitated GluR2 (b) were detected by immunoblotting with anti-GRIP and anti-GluR2 N-terminal antibodies, respectively. Collectively, these results suggest that the disruption of GluR2 clusters by TPA, via PKC phosphorylation of Ser880, results from a loss of anchoring via GRIP
Fig. 5. Disruption of GluR2 clusters concurrently with phosphorylation of the Ser880 residue in GluR2 following KCl + glutamate stimulation. • A cerebellar neuronal culture was stimulated with 50 mM KCl together with 10 µM glutamate for 4 min. Then, the drugs were removed and used for subsequent analysis immediately or after 2 h of incubation. (A) Quantitative analysis of GluR2 cluster intensities. The graph shows the means of the maximal cluster intensities obtained from five different cells fixed immediately or 2 h after KCl + glutamate stimulation (white columns). Black columns show the results from control samples. • (B) Phosphorylation of the Ser880 residue in GluR2 following KCl + glutamate stimulation. Cerebellar neuronal cultures treated (white columns) or not treated (black columns) with KCl + glutamate, and incubated for 0 or 120 min after removal of the drugs. The membrane fractions were subjected to SDS–PAGE and subjected to immunoblotting with anti-P-Ser880 antibodies (insets).
Fig. 6. Internalization of GluR2 protein following PKC activation. • (A) Increase in intensity of immunoreactive intradendritic GluR2. Upper panels show respective confocal optical sections of the dendritic regions in control and TPA-treated Purkinje cells immunostained after permeabilization treatment with anti-GluR2 N-terminal antibodies (scale bar 5 µm). White bars indicate the region whose immunoreactive intensities were quantified. The graph indicates results of the quantitative analysis of intracellular immunoreactive intensities (five cells, total Å`120 µm dendritic length). • (B) Decrease in the amount of extracellularly expressed GluR2 following TPA stimulation. Cell surface proteins were biotinylated followed by immunoprecipitation for GluR2. Then, the biotinylated GluR2 protein was detected by immunoblotting with HRP-conjugated streptavidin. Inset graph indicates the total amount of immunoprecipitated GluR2 protein. (C) Increase in the amount of GluR2 protein contained in the 150 000 g fraction following TPA stimulation. A 150 000 g fraction containing clathrin-coated vesicles was obtained as described in Materials and methods. The amount of GluR2 in this fraction was quantified by immunoblot analysis using anti-GluR2 N-terminal antibodies. The inset graph indicates the results of the quantitative analysis of total GluR2 protein contained in respective cultures.
Fig. 7. A model depicting LTD of AMPA receptor response at parallel fiber–Purkinje cell synapses. • The left part of the figure shows the basal state of the synapse. The intracellular C-terminal tail of GluR2 is bound to GRIP, and thus immobilized at the postsynaptic membrane. The right part of the figure shows the synapse after induction of LTD. Ser880 in the C-terminal domain of GluR2 is phosphorylated, which causes dissociation of GluR2 from GRIP, resulting in a decrease in the density of functional AMPA receptors in the postsynaptic membrane. PSD, postsynaptic density; GRIP, glutamate receptor interacting protein. • The major finding of this work is that stimulations inducing LTD at granule cell–Purkinje cell synapses cause phosphorylation of Ser880 in the C-terminal region of GluR2 and thereby reduce the binding affinity of GluR2 to GRIP, a protein known to be important for AMPA receptor clustering.
Recycling Endosomes Supply AMPA Receptors for LTPM. Park, E. C. Penick, J. G. Edwards, J. A. Kauer, M. D. Ehlers
To test whether transport from recycling endosomes to the plasma membrane was required to maintain the supply of AMPA receptors at the dendritic plasma membrane, we took advantage of a mutant version of the Eps15–homology domain protein EHD1/Rme1 (Rme1-G429R) as well as a constitutively inactive guanosine diphosphate–bound form of the small guanosine triphosphatase (GTPase) Rab11a (Rab11a-S25N). Both of these mutants selectively impair endocytic recycling by preventing trafficking from recycling endosomes to the plasma membrane (fig. S1) • LTP-inducing stimuli increase the number of functional AMPA-type glutamate receptors at the postsynaptic membrane, leading to an increase in AMPA receptor–mediated transmission at excitatory synapses . However, the source of AMPA receptors mobilized for LTP is unknown. • AMPA receptors undergo continuous cycling into and out of the postsynaptic membrane. Upon internalization, AMPA receptors are sorted in early endosomes either to a specialized recycling endosome compartment for reinsertion to the plasma membrane or to late endosomes and lysosomes for degradation (fig. S1)
Fig. 1. Transport through recycling endosomes maintains AMPA receptors at synapses. • (A) The ratio of surface to total endogenous GluR1 on individual hippocampal neurons expressing the indicated constructs • (B) Inhibitory mutants of Rme1 and Rab11a retard AMPA receptor recycling. Live antibody feeding and receptor recycling assays were performed on neurons cotransfected with HA/T-GluR1 and either GFP, GFP-Rme1-G429R, or GFP-Rab11a-S25N (17). Fluorescence signal indicates remaining intracellular HA/T-GluR1 after 0 min and 60 min of recycling. • (C) Means ± SEM of internalized intracellular HA/T-GluR1 on individual neurons from (B) • (D) Overexpressing Rme1-G429R traps internalized AMPA receptors (Int GluR1, green) in recycling endosomes along with internalized Alx-Tf (red). Arrows indicate colocalization.
AMPA receptor recycling assay • Surface HA/T-GluR1 was labeled with mouse anti-HA for 10 min at room temperature and allowed to be internalized for another 10 min. • Surface-remaining HA/T-GluR was blocked with unlabeled mouse secondary antibody (Sigma). • For Figures 1B and C, neurons were incubated for 1 hr at 37°C to allow recycling of internalized receptors. • After fixation, HA/T-GluR1 reinserted into the surface was blocked under nonpermeabilizing conditions with unlabeled mouse secondary antibody . • Neurons were then permeabilized and incubated with Cy3-conjugated secondary antibody to label the remaining intracellular internalized HA/T-GluR1.
Fig. 2. Test whether AMPA receptors recruited during LTP use the transport pathway from recycling endosomes to arrive at synapses. • We used a cell culture model of LTP employing glycine stimulation to activate synaptic NMDA receptors together with selective detection of newly inserted AMPA receptors using HA/T-GluR1. • As expected, 20 to 25 min after a brief application of glycine (200 µM for 3 min), newly inserted AMPA receptors were significantly increased compared to unstimulated control cells (Fig. 2, A and B). This increase was blocked by coapplication of the NMDA receptor antagonist D-AP5 (Fig. 2, A and B) • Hippocampal neurons expressing Rme1-G429R and Rab11a-S25N to prevent transport from recycling endosomes failed to exhibit a detectable increase in AMPA receptor insertion upon glycine stimulation (Fig. 2B). In contrast, neurons expressing wild-type versions of Rme1 and Rab11a exhibited robust glycine-induced AMPA receptor insertion (Fig. 2B). Expression of Rab6a-T27N, a dominant negative version of the Golgi-associated Rab family GTPase Rab6a (21–23), had no effect on glycine-induced AMPA receptor insertion (Fig. 2B) • (A) After a 5-min thrombin treatment to remove HA tags from surface HA/T-GluR1, neurons were stimulated with glycine with or without AP5 and incubated to allow new receptor insertion. Newly inserted HA/T-GluR1 was detected by anti-HA surface labeling. • (B) Quantitative analysis of AMPAR insertion in neurons expressing the indicated constructs.
To determine whether AMPA receptors inserted during LTP derive originally from the plasma membrane or are newly synthesized, the authors tested the effect of protein synthesis inhibition. • Inhibiting protein synthesis in hippocampal neurons (20 µM anisomycin for 4 hours) had no effect on glycine-induced insertion of HA/T-GluR1 (Gly/basal insertion: untreated, 156 ± 16% and n = 37; anisomycin, 144 ± 13% and n = 21; P = 0.63), consistent with the notion that mobilized AMPA receptors were not newly synthesized. • Next, they tested whether previously endocytosed AMPA receptors were delivered to the plasma membrane in response to LTP-inducing stimuli. With use of live-cell antibody feeding, they visualized internalized HA/T-GluR1receptors and followed the loss of intracellular HA/T-GluR1 fluorescence as a direct measure of AMPA receptor recycling back to the plasma membrane. • Under these conditions, internalized AMPA receptors recycled to the plasma membrane, and this recycling was enhanced twofold by glycine stimulation (Fig. 2C, top). The loss of internalized receptors was not due to degradation, because the sum total of intracellular plus recycled AMPA receptors was constant under all conditions (Fig. 2C, bottom).
Fig. 2. Recycling endosomes supply newly inserted AMPA receptors during LTP. • After the standard 5-min thrombin treatment (19) (Fig. 2A), glycine stimulation caused a robust insertion of HA/T-GluR1 at the dendritic plasma membrane (Fig. 2, D and E). In contrast, after prolonged thrombin incubation (4 hours at 37°C) to remove HA tags from all HA/T-GluR1 receptors cycling to the plasma membrane during this time period, glycine-induced insertion of HA/T-GluR1 was undetectable (Fig. 2, D and E). Thus, AMPA receptors inserted during LTP derived from a pool that was present at the plasma membrane in the previous 4 hours. • In neurons expressing Rme1-G429R, Rab11a-S25N, or Syn13TM, depletion of the cycling pool of HA/T-GluR1 had no effect on AMPA receptor insertion (Fig. 2E), establishing the specificity of these reagents for disrupting endocytic recycling but not de novo insertion of newly synthesized receptors. Thus, AMPA receptors mobilized during LTP-inducing stimuli derive from a recycled pool.
Fig. 3. Blocking transport through recycling endosomes abolishes LTP. • (A) Hippocampal slices were infected with Sindbis virus expressing GFP (n = 17), GFP-Rme1-G429R (n = 8), or GFP-Rab11a-S25N (n = 7). EPSCs were measured by whole-cell patch-clamp recordings from GFP-positive CA1 pyramidal neurons before and after HFS (arrow) to induce LTP (100 Hz, four 1-s trains). (B) EPSCs were recorded from CA1 pyramidal neurons with or without an LTP-inducing HFS (arrow). Purified syntaxin13TM (Syn13TM; n is 8 for HFS and 6 for no HFS) or syntaxin7TM (Syn7TM, n = 7) proteins (2 µg/ml each) were included in the recording pipette solution.
Schematic model for the mobilization of AMPA receptors during LTP. • Ongoing AMPA receptor cycling is supported by transport through endosomal compartments. Following NMDA receptor activation, AMPA receptors rapidly exit recycling endosomes and traffic to the postsynaptic membrane, likely through vesicle intermediates. PSD, postsynaptic density; EZ, endocytic zone; EE, early endosome; RE, recycling endosome.
Homeostatic plasticity and postsynaptic receptor trafficking • Alterations in synaptic strength can be encoded presynaptically as alterations in the machinery releasing the neurotransmitter glutamate, or postsynaptically by changing the number or function of receptors sensing the glutamate signal. • A key advance has been the discovery that glutamate receptors are dynamically transported to and from the postsynaptic membrane. Such dynamic transport operates over a wide range of timescales and responds to diverse stimuli, matching the varied spectrum of plasticity modes. • It is now widely accepted that the trafficking of AMPA-type glutamate receptors mediates rapid synaptic modification, long term potentiation (LTP) and long term depression (LTD). • Synaptic scaling is a homeostatic form of plasticity that tends to restore neurotransmission activity to ‘baseline’ levels. • NMDA receptor scaling, as for that of AMPA receptors, occurs through changes in the number of receptors clustered at the synapse.
Deletion of the last 4 amino acids of GluR2, which should abolish interaction with PDZ proteins GRIP/ABP and PICK-1, had only a modest negative effect on steady-state surface level of HA/T-GluR24 (Fig. 7a and b). • The intracellular levels of HA/T-GluR2 4 and wild type were not significantly different. In the thrombin cleavage assay, the time course of surface accumulation of HA/T-GluR2 4 ( = 18 min) was slower than wild type (Fig. 7c). Truncation of the last 15 amino acids of GluR2 (HA/T-GluR215) reduced the steady-state surface expression further (Fig. 7a and b), though the effect was still not as dramatic as with the 4-residue C-terminal truncation of GluR1 (Fig. 6).
Fig. 6.2AR trafficking with Rab4 activation with Q72L (Upper) and inhibition with S27N (Lower). • In both panels, activated sarcolemmal AR are coupled to Gs but translocate to caveolae where they disassociate from G proteins. Receptor internalization, presumably mediated by Rab5 (28), brings AR to sorting vesicles (Upper), where Rab4 promotes their reinsertion into sarcolemma. Rab4 activation accelerates this process, constantly renewing the supply of functional receptors (Upper). Rab4 inhibition (Lower) impairs recycling, causing accumulation of uncoupled receptors in vesicles and caveolae.
Rab4 inhibition alone prevented resensitization after isoproterenol-induced in vivo adrenergic desensitization. Confocal and ultrastructural analyses revealed bizarre vesicular structures and abnormal accumulation of 2AR in the sarcoplasm and subsarcollema of Rab4 S27N, but not Q72L, mice. These data provide evidence for constant bidirectional sarcollemal–vesicular 2AR trafficking in the in vivo heart and show that Rab4-mediated recycling of internalized 2AR is necessary for normal cardiac catecholamine responsiveness and resensitization after agonist exposure. • Fig. 4. (A) Distribution of h-2AR (green fluorescence) in h-2AR and h-2AR/Rab4 S27N compound transgenic myocardia. Arrow indicates granular cytoplasmic staining; arrowhead shows the coarse labeling of the sarcolemma. Red fluorescence is f-actin. Scale bar, 20 µm (Magnification, x600.) (B) Ultrastructural features: h-2AR/Rab4 S27N vesicular structures are intensely convoluted and the membranous profiles abnormally fused.
The EMBO Journal (2005) 24, 2265–2283, doi: 10.1038/sj.emboj.7600688Published online 9 June 2005Essential role of Hrs in a recycling mechanism mediating functional resensitization of cell signalingAylin C Hanyaloglu, Emma McCullagh and Mark von Zastrow • Hepatocyte growth factor-regulated tyrosine kinase substrate (Hrs) is well known to terminate cell signaling by sorting activated receptors to the MVB/lysosomal pathway. Here we identify a distinct role of Hrs in promoting rapid recycling of endocytosed signaling receptors to the plasma membrane. • This function of Hrs is specific for receptors that recycle in a sequence-directed manner, in contrast to default recycling by bulk membrane flow, and is distinguishable in several ways from previously identified membrane-trafficking functions of Hrs/Vps27p. In particular, Hrs function in sequence-directed recycling does not require other mammalian Class E gene products involved in MVB/lysosomal sorting, nor is receptor ubiquitination required. • Disrupting Hrs-dependent recycling prevented functional resensitization of the 2-adrenergic receptor, converting the temporal profile of cell signaling by this prototypic G protein-coupled receptor from sustained to transient. • These studies identify a novel function of Hrs in a cargo-specific recycling mechanism, which is critical to controlling functional activity of the largest known family of signaling receptors.