Download

1 / 39
410 likes | 550 Views
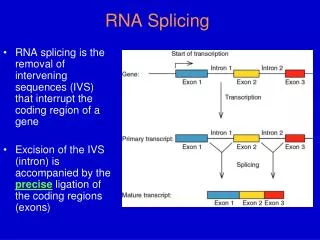
RNA Splicing and Processing. Introduction. Typical mammalian gene has 7-8 exons over ~16 kb Exons are short (~100-200 bp) Introns are long (~>1kb) Pre-mRNA mRNA Splicing occurs in the nucleus. Figure 26.2. Splicing systems --Splicing apparatus --Autonomously

E N D
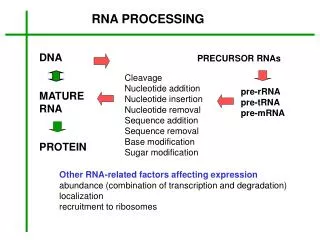
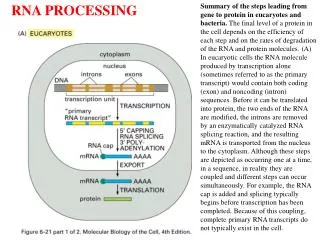
Introduction Typical mammalian gene has 7-8 exons over ~16 kb Exons are short (~100-200 bp) Introns are long (~>1kb) Pre-mRNA mRNA Splicing occurs in the nucleus Figure 26.2 Splicing systems --Splicing apparatus --Autonomously (Self-governing //Independent -Transterification-) --Enzymatic activities (enzymes for RNA) Eukaryotic genes often contain intervening sequencings (introns) that separate the exons and must be removed for proper protein translation to occur. Removal of introns involves a complicated “molecular device” called a spliceosome that involves protein and RNA (like a ribosome). Splicing allows an increase in the information content of a genome due to alternative splicing products that produce novel proteins.
Nuclear Splice Junctions Are Short Sequences • Splice sites are the sequences immediately surrounding the exon–intron boundaries. • They are named for their positions relative to the intron.
Nuclear splice junctions are interchangeable but are read in pairs • The 5′ splice site at the 5′ (left) end of the intron includes the consensus sequence GU. • The 3′ splice site at the 3′ (right) end of the intron includes the consensus sequence AG. • The GU-AG rule describes the requirement for these constant dinucleotides at the first two and last two positions of introns in pre-mRNAs. Splice sites are generic. GT…AG (DNA)-(~98 %);(~2% ?) Directional (one direction) The apparatus for splicing is not tissue specific (with some exceptions). The mechanism by which pairs of splice sites are correctly identified is unknown. Figure 26.3
Splice Junctions Are Read in Pairs Two hypothesis for splicing both 5’ and 3’ pairs: --Intrinsic property of the RNA for matching the 5’ and 3’ region of the intron. --All 5’ and 3’ are functionally equivalents. • Splicing depends only on recognition of pairs of splice junctions. • All 5′ splice sites are functionally equivalent, and all 3′ splice sites are functionally equivalent. Figure 26.4 It does not proceed sequentially order…… It proceed in a preferred order. (unique pathway)
Pre-mRNA Splicing Proceeds through a Lariat (A rope in the form of a lasso) • Splicing requires the 5′ and 3′ splice sites and a branch site just upstream of the 3′ splice site. • The branch sequence is conserved in yeast but less well conserved in higher eukaryotes. Only the splice site sequence and the branch site “A” are required for correct splicing. Introns can be of variable length and sequences. Thus, intron conformation is probably not important.
A lariat is formed when: • the intron is cleaved at the 5′ splice site • the 5′ end is joined to a 2′ position at an “A”at the branch site in the intron • The intron is released as a lariat when: • it is cleaved at the 3′ splice site • the left and right exons are then ligated together Figure 26.6
Nuclear splicing involves trans-esterification GU…UACUAAC….AG “Branch site” Does splicing require energy? Does splicing ever occur in DNA? Figure 26.7
snRNAs Are Required for Splicing • The five snRNPs involved in splicing are U1, U2, U4, U5, and U6. • Together with some additional proteins, the snRNPs form the • spliceosome. The spliceosome is a “molecular device (particle)”that involves proteins and RNAs (small nuclear RNAs) Eukaryotic introns, splicing is done in a series of reactions which are catalyzed by the spliceosome, but there are also self-splicing introns Figure 26.8
All the snRNPs except U6 contain a conserved sequence that binds Sm proteins. • The Sm proteins are recognized by antibodies generated in autoimmune disease (systemic lupus erythematosus -SLE-)
U1 snRNP Initiates Splicing • U1 snRNP initiates splicing by binding to the 5′ splice site by means of an RNA–RNA pairing reaction. The spliceosome assembles at the splice site. It involves proteins and snRNAs (like U1 to U6). Figure 26.9
(SRp) SRp Splicing proceeds through discrete stages Two ways…… Early stages of splicing U2 Intron and exon definition: Serine rich region protein (SRp) 5’--U1 snRNA binding --ASF/SF2 3’--U2AF proteins Linker--SF1/BBP E complex What is the source of Energy?
The E Complex Can be Formed by Intron Definition or Exon Definition • The direct way of forming an E complex is for: • U1 snRNP to bind at the 5′ splice site • U2AF to bind at a pyrimidine tract between the branch site and the 3′ splice site • Another possibility is for the complex to form between: • U2AF at the pyrimidine tract • U1 snRNP at a downstream 5′ splice site
The E complex is converted to the A complex when U2 snRNP binds at the branch site. • If an E complex forms using a downstream 5′ splice site, this splice site is replaced by the appropriate upstream 5′ splice site when the E complex is converted to the A complex. • Weak 3′ splice sites may require a splicing enhancer located in the exon downstream to bind SR proteins directly.
26.8 5 snRNPs Form the Spliceosome • Binding of U5 and U4/U6 snRNPs converts the A complex to the B1 spliceosome. • The B1 spliceosome contains all the components necessary for splicing. • The spliceosome passes through a series of further complexes as splicing proceeds.
Nucleus Cytoplasm Figure 26.13 • Release of U1 snRNP: • allows U6 snRNA to interact with the 5′ splice site • converts the B1 spliceosome to the B2 spliceosome • When U4 dissociates from U6 snRNP, U6 snRNA can pair with U2 snRNA to form the catalytic active site. ?
An Alternative Splicing Apparatus Uses Different snRNPs • An alternative splicing pathway uses another set of snRNPs that comprise the U12 spliceosome. • The target introns are defined by longer consensus sequences at the splice junctions. • They usually include the same GU-AG junctions. • Some introns have the splice junctions AU-AC, including: • U11-dependent • U12-dependent
Alternative splicing involves differential use of splice junctions and more than one mRNA sequences are generated GT…AG (DNA) (~98 %) CG…AG AT…AC (AU…AC) T or t antigen and the various E1A proteins are made in all the cells where the genes are expressed. The ratio of T to t made depends on the concentration of ASF/SF2, a splicing factor (~2% ?) U11_U12 snRNAs (Related or similar to U1_U2) Mechanisms ? Arginine-rich/Serine splicing factor
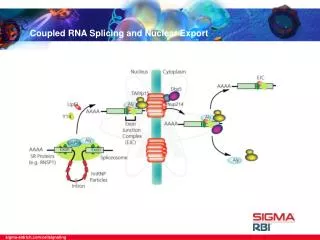
Splicing Is Connected to Export of mRNA • The REF proteins bind to splicing junctions by associating with the spliceosome. • After splicing, they remain attached to the RNA at the exon–exon junction. Figure 26.16 Protein complex: Exon junction complex (EJC)--RNA export factors (REF)
REF proteins interact with the transport protein TAP/Mex. • TAP/Mex exports the RNA through the nuclear pore. Protein complex: Exon junction complex (EJC) Figure 26.17
Mechanisms of Nuclear Import and Export Gene Expression in Prokaryotes: DNA RNA Protein Gene Expression in Eukaryotes: DNA RNAN ----- RNAC Protein export CYTOPLASM --------- Nuclear Proteins Shuttling Proteins NUCLEAR IMPORT/EXPORT ARE ESSENTIAL PROCESSES FOR GENE REGULATION IN EUKARYOTES AND ARE HIGHLY REGULATED PROCESSES N E NUCLEUS --------- Import Export
Nuclear Import Mechanisms • Proteins to be imported into the nucleus have Nuclear Localization Signals (NLS’s) that enable nuclear import. • NLS’s bind to importin α subunit of an importin α-β complex. • Transport through the NPC is mediated by interaction of degenerative sequences in the NPC proteins with the importin β subunit. • Key to function and regulation are RAN GTP [high in nucleus by RCC1 (Ran nucleotide exchange factor)] & RAN GDP [high in cytoplasm by RAN GAP (RAN GTP activating protein)]. • The asymmetric distribution of RCC1 in the nucleus and RAN GAP in the cytoplasm drives the nuclear import process.
NUCLEAR EXPORT • The exporting proteins have special sequences called Nuclear Export Signals (NES’s) that mediate export through binding to a class of proteins that function in export called exportins. • Exportins are typically monomeric and function in a reverse manner to importin under the control of RAN. • Thus the cargo complex requires RAN-GTP which is found only in the nucleus. • Disassociation of the ‘cargo” from the exportin requires RAN-GDP which occurs only in the cytoplasm.
Machinery of nuclear import/export… • RNA Export: • In the current model, RNA export occurs by the export of multiple RNP proteins that cover the mRNA during transport. • So a more precise term is: “RNP (ribonucleoprotein) export”. • In the cell nucleus pre-mRNA is packaged into particles called pre-mRNP or hnRNP particles. protein RNA pre-mRNA RNP particle HnRNP: Heterogeneous nuclear ribonucleoproteins
Summary of Nuclear Splicing --Splicing removes introns from eukaryotic nuclear pre-mRNA. --Most splicing is cis, although trans splicing does occur in the trypanosome (parasite). --Nuclear pre-mRNA splicing is involves the spliceosome made up of proteins and snRNAs.
The 3′ Ends of RNAs pol-I and pol-III Transcripts Are Generated by Termination • RNA polymerase I terminates transcription at an 18-base terminator sequence. • RNA polymerase III terminates transcription in poly(U)4 sequence embedded in a G-C-rich sequence. Figure 26.32
The 3′ Ends of mRNAs Are Generated by Cleavage and Polyadenylation (Pol II) • The sequence AAUAAA is a signal for cleavage to generate a 3′ end of mRNA that is polyadenylated. • The reaction requires a protein complex that contains: • a specificity factor • an endonuclease • poly(A) polymerase Figure 26.34
Endonuclease Poly(A) polymerase Poly(A) binding protein
The specificity factor and endonuclease cleave RNA downstream of AAUAAA. • The specificity factor and poly(A) polymerase add ∼200 A residues progressively to the 3′ end. • A-U-rich sequences in the 3’ tail control cytoplasmic polyadenylation.
Riboswitches A Common RNA Regulatory Element
mRNAs directly sense small-molecule metabolite concentrations. Riboswitch is a regulatory segment of a messenger RNA molecule that binds a small molecule, changing the 3D structure of the mRNA and as consequence resulting in a change in production of the proteins encoded by the mRNA.
---Rho-independent transcriptional terminator (UUUU) with A-U and CG base pairs ---Ribosome-binding site (called the Shine-Dalgarno sequence)