Download
1 / 9

E N D
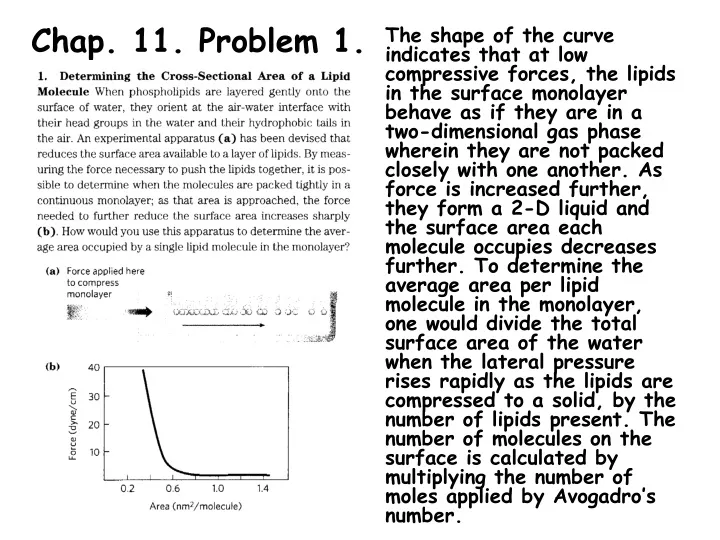
Chap. 11. Problem 1. The shape of the curve indicates that at low compressive forces, the lipids in the surface monolayer behave as if they are in a two-dimensional gas phase wherein they are not packed closely with one another. As force is increased further, they form a 2-D liquid and the surface area each molecule occupies decreases further. To determine the average area per lipid molecule in the monolayer, one would divide the total surface area of the water when the lateral pressure rises rapidly as the lipids are compressed to a solid, by the number of lipids present. The number of molecules on the surface is calculated by multiplying the number of moles applied by Avogadro’s number.
Chap. 11. Problem 3. The molecular weight of an SDS molecule is 288. Thus, if the average molecular weight of a micelle is 18,000, then there are 18,000/288 = 63 SDS molecules per micelle.
Chap. 11. Problem 7. To solve this problem, ignoring the transmembrane electrical potential, the equation ∆Gt = 2.303 RT log (C2/C1) is used. Given that the pH of blood plasma is pH = 7.4 and the pH of gastric acid is pH = 1.5, the H+ concentrations in the two compartments can be calculated using [H+] = 10-pH, which gives Blood plasma [H+] = 10-7.4 = 4.0 x 10-8 M Gastric acid [H+] = 10-1.5 = 3.2 x 10-2 M. Then substituting into the upper equation gives ∆Gt = 2.303 RT log (3.2 x 10-2/4.0 x 10-8) = 35 kJ/mol. Thus the amount of ATP needed for H+ transport into 1 L of gastric juice is 35 kJ/58 kJ/mol = 0.6 mol
Chap. 11. Problem 8. The transport described is against the electrochemical potential across the membrane. Because an ion is being transported, the proper equation for this calculation is ∆Gt = 2.303 RT log (C2/C1) + ZF∆ Because Na+ is being transported to the more positively charged side of the membrane, a positive value of +0.07 V is substituted into the right term so that ∆G > 0 for this term. ∆Gt = 2.303 RT log (145/12) + (1)(96,480 J/Vmol)(0.07 V) ∆Gt = 6.4 kJ/mol + 6.8 kJ/mol = 13 kJ/mol
Chap. 11. Problem 9. The kidney is a highly aerobic tissue that utilizes mitochondrial oxidative phosphorylation to make ATP. In this process, electrons removed in the oxidation of fuel substrates such as glucose or fatty acids are passed to O2 releasing energy that is used to make ATP by the F1Fo ATPase (ATP synthase). If ouabain reduces O2 consumption by 66%, then ATP hydrolyzed by the Na+K+ ATPase pump must account for 66% of the total energy consumption of the kidney.
Chap. 11. Problem 11. Membrane protein topology is commonly assayed by proteolysis. If a membrane protein cannot be cleaved by a protease added from the outside of the intact membrane containing it, then it is likely located on the other side of the bilayer. Because protein X is not cleaved by proteases unless the red blood cell membrane has first been disrupted, the data indicate that the protein is located inside the cell. Because protein X can be removed from the membrane by salt treatment, the combined data indicate that it is a peripheral membrane protein that attaches to the inner leaflet of the RBC plasma membrane.
Chap. 11. Problem 13. The temperature of body extremities is slightly less than the temperature of core tissues. Reindeer, which live in cold climates around the world, maintain the fluidity of the membranes in their extremities by increasing their contents of unsaturated fatty acids in these membrane structural lipids. This compositional change ensures proper membrane dynamics and function in the extremities.
Chap. 11. Problem 19. The data indicate that the leucine transporter has nearly equal affinities (Kts) for L-leucine and L-valine. Therefore, it probably has a hydrophobic binding pocket that can accommodate the side chains of both amino acids, which are similar in structure. The data also indicate that the transporter prefers the L-stereoisomer of leucine, and presumably, valine. Based on the much lower Vmax for transport in the absence of Na+, it is likely that the transporter takes up the amino acids and Na+ via a symport process. Because the Na+K+ ATPase maintains the Na+ gradient across intestinal epithelial cells, transport of L-leucine should be inhibited by ouabain, an inhibitor of the ATPase.
Chap. 11. Problem 22. This 18 residue helix is clearly amphipathic with nonpolar (N) residues on one face and polar residues (P) on the other. When clustered together with other similarly amphipathic helices, the helix could orient with its nonpolar side facing out towards membrane lipids, and its polar side facing other helices in the interior of the protein.