Download
1 / 102
1.23k likes | 1.75k Views
Intracellular compartments Protein Sorting and Transport. Compartments of an Animal Cell. Compartments of an Animal Cell - TEM. Procaryotic Cell. Eucaryotic Cell. Typical Plant Cell. Golgi body. central vacuole. vesicle. rough endoplasm reticulum (rough ER). microfilaments

E N D
Intracellularcompartments Protein Sorting and Transport
Procaryotic Cell Eucaryotic Cell
Typical Plant Cell Golgi body central vacuole vesicle rough endoplasm reticulum (rough ER) microfilaments (components of cytoskeleton) ribosomes (attached to rough ER) ribosomes (free in cytoplasm) smooth endoplasmic reticulum (smooth ER) mitochondrion DNA + nucleoplasm chloroplast nucleolus NUCLEUS nuclear envelope plasma membrane cell wall microtubules (components of cytoplasm)
Plant Cell Animal Cell
Functions of major intracellular compartments: • Nucleus - contains main genome, DNA and RNA synthesis. • Cytosol - most protein synthesis, glycolysis and metabolic pathways synthesizing amino acids, nucleotides. • Endoplasmic reticulum - synthesis of membrane proteins, lipid synthesis. • Golgi apparatus - covalent modification of proteins from ER, sorting of proteins for transport to other parts of the cell. • Mitochondria and chloroplasts (plants) - ATP synthesis. • Lysosomes - degradation of defunct intracellular organelles and material taken in from the outside of the cell by endocytosis. • Endosomes - sorts proteins received from both the endocytic pathway and from the Golgi apparatus. • Peroxisomes - oxidize a variety of small molecules.
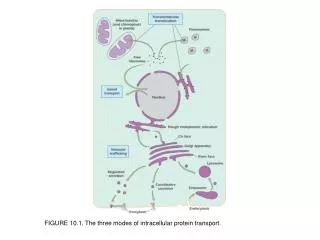
Three basic modes of transport • Gated transport • Transmembrane transport • Vesicular transport
Roadmap of protein traffic The genesis and function of internal compartments depends on the appropriate targeting of proteins Some of the green route illustrated.
A simple experiment shows that many sorting signals consist of a continuous stretch of amino acid sequence called a “signal sequence”. Fusing sorting signals to GFP is particularly good way to do this experiment.
Actin-GFP PAX-GFP
NuclearImport andExport • Nuclear envelope consists of two concentric lipid bilayers. • The perinuclear space is contiguous with the lumen of the ER. • Bidirectional transport occurs through the nuclear pore complexes.
Nuclear Pore The nuclear pore complex is an aqueous channel that allows diffusion of small molecules and proteins up to 60kD. Hence, transport of such molecules is passive.
Molecules larger than 9 nm or 60 KDa must have Nuclear Localization Signals (NLS)and be actively transported into the nucleus
In most cases, nuclear localization of large proteins relies on a signal sequence called a Nuclear localization signal or NLS. • NLS can be located anywhere in the primary sequence of the protein. • Usually arginine and lysine-rich and quite short. • There are some exceptions where nuclear localization relies on a signal patch.
The NLS directs the protein for transport through the nuclear pore complex, and proteins maintain their tertiary and quaternary structures during transport. When gold beads are coated with the NLS, the beads can be seen passing through nuclear pore complexes. The maximum size bead that can be transported is 26 nm. Since the gold bead can’t compress, the opening of the pore must be able to expand.
An asymmetric distribution of a Ran-GAP and a Ran-GEF controls the activity of Ran in a way that allows Ran to mediate active transport through the nuclear pore complex. Ran-GAP in the cytosol causes Ran-GDP to predominate in the cytosol. Ran-GEF in the nucleus causes Ran-GTP to predominate in the nucleus. Ran: Monomeric GTPase Ran-GAP: GTPase-activating protein Ran-GEF: Guanine exchange factor
The NLS associates with soluble cytosolic proteins called nuclear import receptors. The nuclear import receptors also bind the nuclear pore complex so they serve to bring the protein containing the NLS to the nuclear pore complex.
Structure of Ran-GTP & How its binding to import receptor leads to release of cargo protein
Import process 1. Nuclear import receptor associates with cargo and brings the cargo to the nuclear pore. 2. Somehow the receptor/cargo complex moves through the pore (FG-repeats?). 3. Once in the nucleus, Ran-GTP displaces the cargo from the receptor. Ran-GTP is present in the nucleus because of the Ran-GEF. 4. Receptor/Ran-GTP complex move through the pore. 5. Once the receptor returns to the cytosol, Ran-GAP and a Ran binding protein collaborate to cause Ran to dissociate from the receptor and hydrolyze GTP.
The nuclear import players: • Nuclear import receptor binds the cargo. • NLS is in the amino acid sequence of the cargo protein. • Ran-GTP and Ran-GDP are different forms of Ran bound either to GTP or GDP. Ran-GTP causes the NLS to dissociate from the Nuclear import receptor. • Ran-GAP is distinct from Ran but causes Ran to hydrolyze GTP. Hence, Ran-GAP promotes the conversion of Ran-GTP to Ran-GDP. • Ran-GEF is distinct from Ran but causes Ran to release GDP and bind a different molecule of GTP. Hence, Ran-GEF promotes the conversion of Ran-GDP to Ran-GTP.
Export of RNA and proteins relies on nuclear export receptors associating with nuclear export signals (NES) found on proteins and RNA-bound proteins. • Nuclear export receptors are structurally similar to nuclear import receptors. • Nuclear export receptor bind the nuclear export signals and bring the protein to the nuclear pore complex for subsequent transport. • Transport occurs through the same pores through which proteins are imported from the cytosol. • Ran regulates the interaction between the export receptor and the “NES”. The Ran-GTP promotes association of the receptor/cargo complex with the pore in the nucleus and hydrolysis of the GTP on the cytosolic side causes the resulting Ran GDP to dissociate the export receptor from its cargo. • Note - nuclear export receptors do not bind directly to RNA, they bind proteins bound to the RNA.
Transport of ProteinsintoMitochondriaandChloroplasts • Organelles specialized for ATP synthesis. • Most, but not all, proteins are encoded by the nuclear genome and synthesized in the cytosol. • Proteins must be transported to one of multiple membranes or compartments.
Mit. Import signal Polar (+) and Protein Transporters of the Mit. non-polar striped membranes alpha-helix
Import of the protein into the matrix is directed by an N-terminal signal sequence. • For polypeptides encoded by the nuclear genome, synthesis of the polypeptide is first completed in the cytosol. Transport occurs by a posttranslational mechanism. • Signal sequence at the N-terminus associates with the TOM complex located in the outer mitochondrial membrane. TOM is both a receptor for the signal sequence and a translocase. • The polypeptide is passed from TOM to TIM in the innermembrane. During the transport process, the polypeptide traverses both inner and outer membranes via the two translocators at a point known as a contact site. • The polypeptide is imported and the signal peptide is removed by a signal peptidase.
The energetics of import - import requires ATP hydrolysis and an electrochemical proton gradient. • ATP hydrolysis regulates the association of chaperone proteins in the cytosol that serve to keep the polypeptide in an unfolded state prior to association with TOM. • Electrochemical proton gradient in the inner membrane draws the signal sequence through TIM into the matrix. • Chaperone proteins in the matrix use the energy of ATP hydrolysis to pull the polypeptide into the matrix and guide proper folding.
The N-terminal signal sequence for mitochondrial import is a positively charged, amphipathic alpha helix. Hence, the membrane potential across the inner mitochondrial membrane may “electrophorese” the signal sequence through the TIM complex.
Protein import into the inner mitochondrial membrane or intermembrane space
Transport into chloroplasts is similar to mitochondria except there is a 3rd membrane that can be targeted. Targeting the thylakoid membrane involves a second signal sequence. In the case of chloroplasts, the electrochemical proton gradient is at the thylakoid membrane where this gradient participates in transport. Transport across the chloroplast inner membrane is powered by GTP and ATP hydrolysis.
Peroxisomes (which are not part of the endomembrane system) contain enzymes used to incorporate hydrogens with oxygen to make hydrogen peroxide which is then converted to water and oxygen. They function in a variety of situations including detoxifying poisons by transferring hydrogens from them to oxygen. The dense bodies within the peroxisomes are crystallized enzymes. Peroxisomes are formed from phospholipids and enzymes in the cytosol, not by pinching off from the endomembrane system.
Peroxisomes are organelles that perform a variety of oxidation reactions including ones that breakdown fatty acids and toxic molecules that enter the cell from the blood stream. Genetic defects in peroxisomes often cause neurological problems because a particular lipid found in myelin is produced in the peroxisomes. Import of proteins involves short signal sequences. The most unusual aspect of the transport process is that oligomeric proteins don’t have to unfold. Very little is known about the transport process.
Rough ER Rough ER’s rough appearance is due to ribosomes. ER lumen is one continuous space that merges with the perinuclear space. Smooth ER
Functions of the ER • Starting point for newly synthesized proteins destined for Golgi, Endosome, Lysosomes, Secretory vesicles, and the Plasma membrane (see below). • Establishes orientation of proteins in the membrane. • Site of phospholipid and cholesterol synthesis. • Initiation site for N-linked glycosylation of proteins. • Sequesters Ca++ - sarcoplasmic reticulum in muscle is a specialized ER.
A signal sequence of approximately 20 amino acids and rich with hydrophobic amino acids is often located at the N-terminus. • Since the ribosome masks about 30 amino acids, the signal sequence isn’t fully exposed until the nascent polypeptide is about 50 amino acids long. • SRP-ribosome attaches to SRP receptor and then docks on a protein translocator. • SRP and receptor dissociate. • Translation and translocation proceed in unison - co-translational transport. • The energy for transport is provided by the translation process - as the polypeptide grows, it is pushed through the protein translocator. • SRP: signal-recognition particle • SRP receptor
The signal sequence of secreted proteins is cleaved by a signal peptidase. In the literature, the signal sequence of secreted proteins is often called a “leader peptide”.
Co-translational transport must be able to generate a diverse array of configurations. For both single-pass and multipass transmembrane proteins, some types will have the N-terminus projecting into the cytosol and others will have the C-terminus projecting into the cytosol.