Download

1 / 74
860 likes | 1.34k Views
Chapter 13 Gene Regulation in Eukaryotes. Eukaryotic gene regulation occurs at several levels. 1- Control at DNA level by DNA methylation. Small percentages of newly synthesized DNAs (~3% in mammals) are chemically modified by methylation.

E N D
Chapter 13 Gene Regulation in Eukaryotes
1- Control at DNA level by DNA methylation • Small percentages of newly synthesized DNAs (~3% in mammals) are chemically modified by methylation. • Methylation occurs most often in symmetrical CG sequences. • Transcriptionally active genes possess significantly lower levels of methylated DNA than inactive genes. • Methylation results in a human disease called fragile X syndrome; FMR-1 gene is silenced by methylation.
2- Control at DNA level by Histone modifications(Chromatin Remodeling) • Acetylation (乙酰化)by histone acetyl transferases (HATs) and coactivators leads to euchromatin formation; p53 acetylation • Methylationby HDACs (去乙酰化酶)and corepressors leads to heterochromatin formation . Rb-E2F
3-Control at DNA level by gene amplification Repeated rounds of DNA replication yield multiple copies of a particular chromosomal region.
4- Control at transcription initiation By using different sequences (promoter, enhancer or silencer sequences) and factors, the rate of transcription of a gene is controlled gene control region for gene X
5- Control at mRNA splicing (alternate splicing) (four exons) Calcitonin gene-related peptide 61 32 amino acids Reduces bone resorption 37 amino acids Vasodilator
6- Control at mRNA stability • Messenger RNA longevity can be influenced by several factors. • Poly(A) tails seem to stabilize mRNAs. • The sequence of the 3’untranslated region (3’UTR) preceding a poly(A) tail also seems to affect mRNA stability. • Several short-lived mRNAs have the sequence AUUUA repeated several times in their 3’untranslated regions.
6- Control at mRNA stability • When this sequence is artificially transferred to the 3’untranslated region of more stable mRNAs, they, too, become unstable. • Chemical factors, such as hormones, may also affect mRNA stablility. • In the toad Xenopus laevis(非洲爪蟾) , the vitellogenin gene(卵黄生成素) is transcriptionally activated by the steroid hormone estrogen(类固醇激素 ) . However, in addition to inducing transcription of this gene, estrogen also increases the longevity of its mRNA.
6- Control at mRNA stability • Recent research has revealed that the stability of mRNAs and the translation of mRNAs into polypeptides are also regulated by small, noncoding RNA molecules called microRNA (miRNAs).
7- Control at initiation of translation 3’ UTR 5’ UTR AUG UAA Specific sequences make specific secondary structures Specific protein factors bind to these secondary structures
COOH NH NH2 NH2 CO CO NH 8-Regulation by protein stability • Ubiquitin-dependent proteolysis. • Protein molecule is tagged for degradation by attachment of a 20 kDa protein, ubiquitin ATP + 26S proteasome Doomed protein molecule • ubiquitin protein ligase 蛋白酶体系统(ubiquitin-proteasome system(UPS))主要由泛素激活酶(E1)、泛素交联酶(E2)、泛素连接酶(E3)和26S蛋白酶体组成,是降解细胞内蛋白质的主要途径 对于许多细胞进程,包括细胞周期、基因表达的调控、氧化应激反应等,都是必不可少的。2004年诺贝尔化学奖.
Similarity of regulation between eukaryotes and prokaryote 1.Principles are the same: signals (信号), activators and repressors (激活蛋白和阻遏蛋白) recruitment and allostery, cooperative binding (招募,异构和协同结合) 2. The gene expression steps subjected to regulation are similar, and the initiation of transcription is the most pervasively regulated step.
Difference in regulation between eukaryotes and prokaryote***** • Pre-mRNA splicing adds an important step for regulation. (mRNA前体的剪接) • The eukaryotic transcriptional machinery is more elaborate than its bacterial counterpart. (真核转录机器更复杂) • Nucleosomes and their modifiers influence access to genes. (核小体及其修饰体) • Many eukaryotic genes have more regulatory binding sites and are controlled by more regulatory proteins than are bacterial genes. (真核基因有更多结合位点)
A lot more regulator bindings sites in multicellular organisms reflects the more extensive signal integration Bacteria Yeast Human
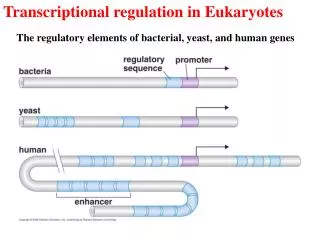
Cis-acting element • Promoter Core promoter in eukaryote: TATA-box, Initiator (Inr) in prokaryote: -10 region, Inr Proximal elements of promoter in prokaryote: -35 region in eukaryote: CAAT-box, GC-box UPE: upstream promoter element UAS: upstream activating sequence • Terminator (终止子):A DNA sequence just downstream of the coding segment of a gene, which is recognized by RNA polymerase as a signal to stop transcription.
Enhancer (激活元件) :a given site binds regulator responsible for activating the gene.Alternative enhancerbinds different groups of regulators and control expression of the same gene at different times and places in responsible to different signals. Activation at a distance is much more common in eukaryotes. Silencer (沉默子)A DNA sequence that helps to reduce or shut off the expression of a nearby gene. Insulators (绝缘子) or boundary elements (临界元件) are regulatory sequences between enhancers and promoters. They block activation of a linked promoter by activator bound at the enhancer, and therefore ensure activators work discriminately.
What is trans-acting factor? Usually they are proteins, that bind to the cis-acting elements to control gene expression.
These trans-acting factors can control gene expression in several ways: may be expressed in a specific tissue may be expressed at specific time in development may be required for protein modification may be activated by ligand binding
(1) RNA polymerase prokaryotic RNA Pol eukaryotic RNA Pol (2) Transcription factors Basal/general TFs Specific TFs
(3) Domains of trans-acting factors DNA binding domain DBD DNA结合结构域 transcription activating domain 转录活化结构域
一、真核的转录激活蛋白的结构特征 The structure features of the eukaryotic transcription activators Topic 1: Conserved Mechanisms of Transcriptional Regulation from Yeast (酵母) to Mammals (哺乳动物)
The basic features of gene regulation are the same in all eukaryotes, because of the similarity in their transcription and nucleosome structure. • Yeast is the most amenable to both genetic and biochemical dissection, and produces much of knowledge of the action of the eukaryotic repressor and activator. • The typical eukaryotic activators works in a manner similar to the simplest bacterial case. • Repressors work in a variety of ways.
1. Eukaryotic activators (真核激活蛋白) have separate DNA binding and activating functions, which are very often on separate domains of the protein.***** Gal4 bound to its site on DNA
Eukaryotic activators---Example 1: Gal4***** • Gal4 is the most studied eukaryotic activator • Gal4 activates transcription of the galactose genes in the yeast S. cerevisae. • Gal4 binds to four sites (UASG) upstream of GAL1(5'-CGGRNNRCYNYNYNCNCCG-3'), and activates transcription 1,000-fold in the presence of galactose The regulatory sequences of the Yeast GAL1 gene.
Experimental evidences showing that Gal4 contains separate DNA binding and activating domains. • Expression of the N-terminal region (DNA-binding domain) of the activator produces a protein bound to the DNA normally but did not activate transcription. • Fusion of the C-terminal region (activation domain) of the activator to the DNA binding domain of a bacterial repressor, LexA activates the transcription of the reporter gene. Domain swap experiment
Domain swap experiment***** Moving domains among proteins, proving that domains can be dissected into separate parts of the proteins. Many similar experiments shows that DNA binding domains and activating regions are separable.
Box1 The two hybrid Assay (双杂交) to study protein-protein interaction and identify proteins interacting with a known protein in cells***** Fuse protein A and protein B genes to the DNA binding domain and activating region of Gal4, respectively. Produce fusion proteins
2. Eukaryotic regulators use a range of DNA binding domains, but DNA recognition involves the same principles as found in bacteria.***** • Helix-turn-helix ( HTH) (螺旋-转角-螺旋) • Zinc finger (锌指)and zinc cluster • Leucine zipper motif(亮氨酸拉链) • Helix-Loop-Helix proteins (螺旋-突环-螺旋): basic zipper and HLH proteins Transcription factor motifs
HTH (helix-turn-helix) α-helix (N-terminus)----specific α-helix (C-terminus)----non-specific
Bacterial regulatory proteins • Most use the helix-turn-helix (HTH:旋转-转角-旋转)motif to bind DNA target • Most bind as dimers to DNA sequence: each monomer inserts an a helix into the major groove. • Eukaryotic regulatory proteins • Recognize the DNA using the similar principles, with some variations in detail. • In addition to form homodimers (同源二聚体), some form heterodimers (异源二聚体) to recognize DNA, extending the range of DNA-binding specificity.
Zinc containing DNA-binding domains (锌指结构域):Zinc finger proteins (TFIIIA) and Zinc cluster domain (Gal4)
Leucine Zipper Motif (亮氨酸拉链基序) : The Motif combines dimerization and DNA-binding surfaces within a single structural unit.
3. Activating regions(激活区域)are not well-defined structures*** • The activating regions are grouped on the basis of amino acids content. • Acidic activation region (酸性激活区域): contain both critical acidic amino acids and hydrophobic aa. yeast Gal4 • Glutamine-richregion (谷氨酰胺富集区): mammalian activator SP1 • Proline-rich region (脯氨酸富集区): mammalian activator CTF1
二、真核转录激活蛋白的招募调控方式和远距调控特征二、真核转录激活蛋白的招募调控方式和远距调控特征 Activation of the eukaryotic transcription by recruitment & Activation at a distance Topic 2: Recruitment of Protein Complexes to Genes by Eukaryotic Activators
Eukaryotic activators (真核激活蛋白) also work by recruiting (招募) as in bacteria, but recruit polymerase indirectly in two ways: 1. Interacting with parts of the transcription machinery. 2. Recruiting nucleosome modifiers that alter chromatin in the vicinity of a gene.
1. Activators recruit the transcription machinery to the gene. The eukaryotic transcriptional machinery contains polymerase and numerous proteins being organized to several complexes, such as the Mediator and the TFⅡD complex. Activators interact with one or more of these complexes and recruit them to the gene.
Chromatin Immuno-precipitation (ChIP) (染色质免疫共沉淀)to visualize where a given protein (activator) is bound in the genome of a living cell*****.)
2. Activators also recruit modifiers that help the transcription machinery bind at the promoter • Two types of Nucleosome modifiers : • Those add chemical groups to the tails of histones (在组蛋白尾上加化学基团), such as histone acetyl transferases (HATs) • Those remodel the nucleosomes (重塑核小体), such as the ATP-dependent activity of SWI/SNF.
How the nucleosome modification help activate a gene?***** • “Loosen” the chromatin structure by chromosome remodeling and histone modification such as acetylation, which uncover DNA-binding sites that would otherwise remain inaccessible within the nucleosome.
Local alterations in chromatin structure directed by activators Activators, capable of binding to their sites on DNA within a nucleosome are shown bound upstream of a promoter that is inaccessible within chromain. • The activator is shown recruiting • a histone acetylase. That enzyme • adds acetyl groups to residues within • the histone tails. This alters the • packing of the nucleosomes • somewhat, and also creates binding • sites for proteins carrying the • appropriate recognition domains. (b)The activator recruits a nucleosome remodeller, which alters the structure of nucleosomes around the promoter, rendering it accessible and capable of binding the transcription machinery.