Download

1 / 86
880 likes | 1.01k Views
Chapter 17 Gene Regulation in Eukaryotes. 04 级生物学基地班 姜冠男 200431060045. This chapter can be studied with comparison to chapter 16. F O R E W O R D. The regulation in eukaryotes is very like that in prokaryote, but is more complex.

E N D
Chapter 17 Gene Regulation in Eukaryotes 04级生物学基地班 姜冠男 200431060045
This chapter can be studied with comparison to chapter 16
F O R E W O R D The regulation in eukaryotes is very like that in prokaryote, but is more complex. The basic principles and steps of regulation are similar, but eukaryotic genes have additional steps and more regulatory binding sites and are controlled by more regulatory proteins. Besides, nucleosomes and their modifiers influence access to genes.
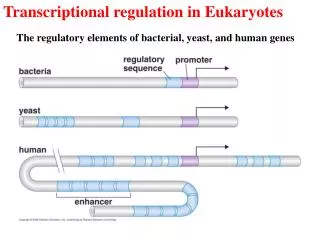
The increasing complexity of regulatory sequences from a simple bacterial gene controlled by a repressor to a human gene controlled by multiple activators and repressors.
O U T L I N E • Conserved Mechanisms of Transcriptional Regulation from Yeast to Mammals • Recruitment of Protein Complexes to Genes by Eukaryotic Activators • Signal Integration and Combinatorial Control • Transcriptional Repressors • Signal Transduction and the Control of Transcriptional Regulators • Gene “Silencing” by Modification of Histones and DNA • Eukaryotic Gene Regulation at Steps after Transcription Initiation • RNAs in Gene Regulation
1CONSERVED MECHANISMS OF TRANSCRIPTIONAL REGULATION FROM YEAST TO MAMMALS
Not all details of gene regulation are the same in all eukaryotes, though they have much in common. A typical yeast gene has less extensive regulatory sequences than its multicellular counterpart. Repressors work in a variety of ways.
ⅠActivators have separate DNA binding and activating functions. The yeast activator Gal4 binds as a dimer to a 17bp site on DNA. The activation domain and the binding domain are separated.
Gal4 binds to four sites located 275bp upstream of Gal1, and activates transcription Of Gal1 gene 1000-fold in the presence of galactose.
The DNA-binding • domain of Gal4, without that • protein’s activation domain, • can still bind DNA, but cannot • activate transcription. (b) Attaching the activation Domain of GA4 to the DNA- binding domain of the bacterial protein LexA, creates a hybrid protein that activates transcription of a gene in yeast as long as that gene bears a binding site for LexA. Domain swap experiment
The Two Hybrid Assay This assay is used to identify proteins that interact with each other.
Ⅱ Eukaryotic Regulators Use a Range of DNA-Binding Domains, but DNA Recognition Involves the Same Principles as Found in Bacteria • Several of the regulatory proteins in eukaryotes bind DNA as heterodimers, and in some cases even as monomers. Heterodimers extend the range of DNA-binding specificities available. • Homeodomain Proteins. The homeodomain is a class of helix-turn-helix DNA-binding domain and recognizes DNA in essentially the same way as those bacterial proteins. • Zinc Containing DNA-Binding Domains. There are various different forms of DNA-binding domain that incorporate a zinc atoms: zinc finger and zinc cluster. • Leucine Zipper Motif. This motif combines dimerization and DNA- binding surfaces within a single structural unit. • Helix-Loop-Helix Proteins. AS in the example of the leucine zipper, an extended ɑ helical region from each of two monomers inserts into the major groove of the DNA.
DNA recognition by a homeodomain. The homeodomain consists of three ɑ helices, of which two form the structure resembling the helix- turn-helix motif. Thus, helix 3 is the recognition helix and is inserted into the major groove of DNA.
Zinc finger domain The ɑ helix on the left of the structure is the recognition helix, and is presented to the DNA by the β sheet on the right. The zinc is coordinated by the two His residues in the ɑ helix and two Cys residues in the β sheets as shown. This arrangement stabilizes the structure and is essential for DNA binding.
Two large ɑ helices, one from each monomer, form both the dimerization and DNA-binding domain at different sections along their length. Thus, as shown, toward the top the two helices interact to form a coiled-coil that holds the monomers together; further down, the helices separate enough to embrace the DNA, inserting into the major groove on opposite sides of the DNA-helix. Once again, specificity is provided by contacts made between amino acid side chains on the ɑ helices and the edge of base pairs in the major groove. zipper bound to DNA.
Helix-loop-helix motif. A long ɑ helix involved in both DNA recognition and , in combination with a second shorter, ɑ helix dimerization.
Ⅲ Activating Regions Are Not Well-Defined Structures • The activating regions are grouped • on the basis of amino acids content • Acidic activation domains • Glutamine-rich domains • Proline-rich domains
2RECRUITMENT OF PROTEIN COMPLEXES TO GENES BY EUKARYOTIC ACTIVATORS
The activator recruits polymerase indirectly in two ways: ⅠActivators Recruit the Transcriptional Machinery to the Gene • 1 . The activator can interact with parts of the • transcription machinery other than • polymerase, and , by recruiting them, recruit • polymerase as well . • 2 . Activators can recruit nucleosome modifiers • that alter chromatin in the vicinity of a gene • and thereby help polymerase bind. • In many cases, a given activator can work in • both ways.
Activation of transcription initiation in eukaryotes by recruitment of the transcription machinery. A single activator is shown recruiting two possible target complexes: the Mediator; and, through that, RNA polymerase Ⅱ; and also the general transcription factor TFIID. Other general transcription factors are recruited as part of the Mediator\ Pol Ⅱ complex; separately, or bind spontaneously in the presence of the recruited components.
Activation of transcription through direct tethering of mediator to DNA The GAL1 gene is activated, in the absence of its usual activator Gal4, by the fusion of the DNA- binding domain of LexA to a component of the Mediator Complex. Activation depends on LexA DNA-binding sites being inserted upstream of the gene. Other components required for transcription initiation presumably bind together with Mediator and Pol Ⅱ.
At most genes, the transcription machinery is not prebound, and appear at the promoter only upon activation. Thus, no allosteric activation of the prebound polymerase has been evident in eukaryotic regulation.
ⅡActivators also Recruit Nucleosome Modifiers that Help the Transcription Machinery Bind at the Promoter • Nucleosome modifiers come in two types: • Those that add chemical groups to the tails of • histone such as histoneacetyl transferases • (HATs) , which add acetyl groups; • Those that remodel the nucleosomes, such as • the ATP-dependent activity of SWI\ SNF.
How do these modifications help activate a gene? • Two basic models to explain how changes in • nucleosomes can help the transcriptional machinery • bind at the promoter: • Remodeling, and certain modifications, • can uncover DNA-binding sites that • would otherwise remain inaccessible • within the nucleosome. • Creating specific binding sites on • nucleosomes for proteinsbearing • so-called bromodomains.
Local alterations in chromatin structure directed by activators Activators, capable of binding to their sites on DNA within a nucleosome are shown bound upstream of a promoter that is inaccessible within chromain. • The activator is shown recruiting • a histone acetylase. That enzyme • adds acetyl groups to residues within • the histone tails. This alters the • packing of the nucleosomes • somewhat, and also creates binding • sites for proteins carrying the • appropriate recognition domains. (b)The activator recruits a nucleosome remodeller, which alters the structure of nucleosomes around the promoter, rendering it accessible and capable of binding the transcription machinery.
Ⅲ Action at a Distance: Loops and Insulators Many eukaryotic activators---particularly in higher eukaryotes---work from a distance. Various models have been proposed to explain how proteins binding in between enhancers and promoters might help activation in the cells of higher eukaryotes. • In Drosophila, the cut gene is activated from an enhancer • some 100 kb away. A protein called Chip aids communication • between enhancer and gene. • In eukaryotes, the DNA is wrapped in nucleosomes, and • the histones within those nucleosomes are subject to • various modifications that affect their disposition and • compactness.
Specific elements called Insulators control the actions of activators, preventing the activating of the non-specific genes.
a) A promoteractivated by activators bound to an enhancer. b) An insulator is placed between the enhancer and the promoter. When bound by appropriate insulator- binding proteins, activation of the promoter by the enhancer is blocked, despite activators binding to the enhancer. c) The activator can activate another promoter nearby. s) The original promoter can be activated by another enhancer placed downstream. Insulators block activation by enhancers.
Transcriptional silencing Silencing is a specialized form of repression that can spread along chromatin, switching off multiple genes without the need for each to bear binding sites for specific repressors. Insulator elements can block this spreading, so insulators protect genes from both indiscriminate activation and repression. Agene inserted at random into the mammalian genome is often “silenced” because incorporated into a particularly dense form of chromatin called heterochromatin. But if insulators are placed up- and downstream of that gene they protect it from silencing.
Ⅳ Appropriate Regulation of Some Groups of Genes Requires Locus Control Regions Only in adult bone marrow are the correct regulators all active and present in appropriate concentrations to bind these enhancers. But more than this is required to switch on these genes in the correct order.
A group of regulatory elements collectively called the locus control region, or LCR, is found 30-50 kb upstream of the whole cluster of globin genes. It binds regulatory proteins that cause the chromatin structure around the whole globin gene cluster to “OPEN UP”, allowing access to the array of regulators that control expression of the individual genes in a defined order.
The human • globin genes and the • LCR that ensures • their ordered • expression. • (b) The globin genes • from mice, which are • also regulated by an • LCR. • (C) The HoxD gene • cluster from the • mouse controlled by • an element called • the GCR which like • the LCRs appears to • impose ordered • expression on the • gene cluster. Regulation by LCRs
3SIGNAL INTERATION AND COMBINATORIAL CONTROL
Ⅰ Activators Work Together Synergistically to Integrate Signals • In multicellular organisms signal integration is used • extensively. In some cases numerous signals are required • to switch a gene on. So at many genes multiple activators • must work together to switch the gene on. • When multiple activators work together, they do so • synergistically. That is , the effect of two activators • working together is greater than the sum of each of them • working alone. • Three strategies of synergy: • Two activators recruit a single complex • Activators help each other binding cooperativity
Cooperative • binding through • direct interaction • between the two proteins. • (b) A similar effect is • achieved by both • proteins interacting with • a common third protein. • (c) The first protein recruits • a nucleosome remodeller • whose action reveals a • binding site for a second • protein. • (d) Binding a protein • unwinds the DNA from • nucleosome a little, r • revealing the binding site • for another protein. Cooperative binding of activators
Ⅱ Signal Integration: the HO Gene Is Controlled by Two Regulators; One Recruits Nucleosome Modifiers and the Other Recruits Mediator. The HO gene is involved in the budding of yeast. The HO gene is expressed only in mother cells and only at a certain point in the cell cycle. These two conditions are communicated to the gene through two activators: SWI5 and SBF.
Why does expression of the gene depend • on both activators? • SBF( which is active only at the correct stage • of the cell cycle) cannot bind its sites unaided; • their disposotion within chromatin prohibits it. • SWI5( Which acts only in the mother cell) can • bind to its sites unaided but cannot, from that • distance, activate the HO gene. • SWI5 can recruit nucleosome modifiers. • These acts on nucleosomes over the SBBF sites. • Thus, if both activators are present and active, • the action of SWI5 enables SBF to bind, and • that activator, in turn, recruits the transcriptional • machinery and activates expression of the gene.
SWI5 can bind its sites within chromatin unaided, but SBF cannot. Remodellers and histone acetylases recruited by SWI5 alter nucleosomes over the SBF sites, allowing that activator to bind near the promoter and activate the gene. Control of the HO gene
Ⅲ Signal Integration: Cooperative Binding of Activators at the Human β-Interferon Gene The human β-interferon gene is activated in cells upon viral infection. Infection triggers three activators: NFĸB, IRF, and Jun\ ATF. They bind cooperatively to sites within an enhancer, form a structure called enhanceosome.
The human β-interferon enhanceosome Cooperative binding of the three activators, to gether with the architectural protein HMG-1, activates the β-interferon gene.
Ⅳ Combinatorial Control Lies at the Heart of the Complesity and Diversity of Eukaryotes There is extensive combinatorial control in eukaryotes. In complex multicellular organisms, combinatorial control involves many more regulators and genes, and repressors as well as activators can be involved.
Combinatorial control Each of the two gene controlled by multiple signals- four in the case of gene A; three in the case of gene B. Each signal is communicated to a gene by one regulatory protein. Regulatory protein 3 acts at both genes, in combination with different additional Regulators in the two cases.
Ⅴ Combinatorial Control of the Mating-Type Genes from Saccharomyces cerevisiae The yeast S. cerevisiae exists in three forms: Two haploed cells of different mating types-a and ɑ-and the diploid formed when and an a and an ɑ cell mate and fuse. Cells of the two mating types differ because they express different sets of gene: a specific genes and ɑ specific genes.
The a cell and the ɑ cell each • encode cell type specific regulators: • a cells make the regulatory protein a1; • ɑ cells make the proteins ɑ1 and ɑ2. • A fourth regulatory protein called Mcm1, • is also involved in regulating the • mating-type specific genes and is • present in both cell types.
The three cell types of the yeast S. cerevisiae are defined by the sets of genes they express. One ubiquitous regulator( Mcm1) and three cell-type specific regulators( a1, ɑ1 and ɑ2) together regulate three classes of target genes. The MAT locus is the region of the genome which encode the mating type regulator. Control of cell-type specific genes in yeast
4 TRANSCRIPTIONAL REPRESSORS
Repressors don’t work by binding to sites that overlap the promoter and thus block binding of polymerase. Another form of repression different from bacteria, which is the most common in eukaryotes. It works as follows: As with activators, repressors can recruit nucleosome modifiers, but in this case the enzymes have the opposite effects to those recruited by activators---they compact the chromatin or remove groups recognized by the transcriptional machinery.
a) By binding to a site on DNA that overlaps the binding site of an activator, a repressor can inhibit binding of the activator to a gene, and thus block activation of that gene. b) A repressor binds to a site on DNA beside an activator and interacts with that activator, occluding its activating region. c) A repressor binds to a site upstream of a gene and, by interacting with the transcriptional machinery at the promoter in some specific way, inhibits transcription initiation. d) repression by recruiting histone modifiers that alter nucleosomes in ways that inhibit transcription. Ways in which eukaryotic repressors work
In the presence of glucose, Mig1 binds a site between the UASG and the GAL1 promoter. By recruiting the Tup1 repressing complex, Mig1 repress expression of GAL1. Repression is a result of deacetylation of local nucleosomes, and also probably by directly contacting and inhibiting the transcription machinery. Repression of the GAL1 gene in yeast
5 SIGNAL TRANSDUCTION AND THE CONTROL OF TRANSCRIPTIONAL REGULATORS