Download
1 / 21
300 likes | 818 Views
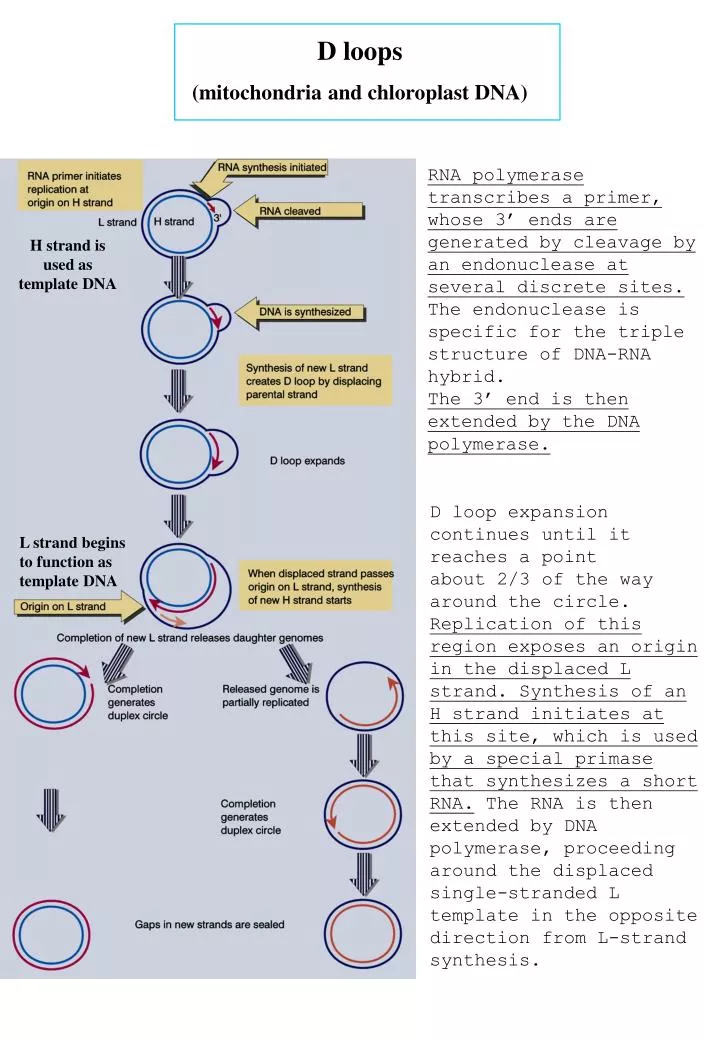
D loops (mitochondria and chloroplast DNA). RNA polymerase transcribes a primer, whose 3’ ends are generated by cleavage by an endonuclease at several discrete sites. The endonuclease is specific for the triple structure of DNA-RNA hybrid. The 3’ end is then extended by the DNA polymerase.

E N D
D loops (mitochondria and chloroplast DNA) RNA polymerase transcribes a primer, whose 3’ ends are generated by cleavage by an endonuclease at several discrete sites. The endonuclease is specific for the triple structure of DNA-RNA hybrid. The 3’ end is then extended by the DNA polymerase. H strand is used as template DNA D loop expansion continues until it reaches a point about 2/3 of the way around the circle. Replication of this region exposes an origin in the displaced L strand. Synthesis of an H strand initiates at this site, which is used by a special primase that synthesizes a short RNA. The RNA is then extended by DNA polymerase, proceeding around the displaced single-stranded L template in the opposite direction from L-strand synthesis. L strand begins to function as template DNA
Rolling circle (phage as M13, f1 and X174). Replicative form Circular duplex DNA (RF) Initiation occurs on one strand. The A protein, nicks the (+) strand of the duplex DNA at a specific site that defines the origin for replication. Elongation of growing strand displaces old strand Continued elongation generates displaced strand of multiple unit lengths Circular ssDNA is packaged into the phage virion (M13, f1, X174). Upon infection single-stranded DNA is converted to double-stranded DNA (RF) (ssDNA) It may be used as a rolling circle to generate more progeny (dsDNA)
Plasmid ColE1 5’ The sequence of RNA I is complementary to the 5’ region of primer RNA origin 5’ 3’-OH 5’ 5’ Proposed model No base pairing between RNA I (not produced) and RNA primer DNA replication RNase H Rop protein stabilizes RNA structure Base pairing with RNA I may change the secondary structure of the primer RNA No DNA replication
The problem of linear replicons - The problem may be circumvented by converting the linear replicon into a circular or multimeric molecule (T4 and lambda phages). -The DNA may form an unusual structure (hairpin). - Instead of being precisely determined, the end may be variable (eukaryotic chromosomes). - The most direct solution is for a protein to intervene to make initiation possible at the actual terminus. Adenovirus 55 K daltons protein Initiation of DNA replication In several viruses that use such mechanisms, a protein is found covalently attached to each 5’end. In the case of adenovirus, a terminal protein is linked to the mature viral DNA via a phosphodiester bond to serine.
Telomeric DNA Preventing the ends of a linear dsDNA molecule from gradually getting shorter during successive rounds of DNA replication. The telomerase extends the telomeric DNA at the 3’-end The telomerase is a reverse transcriptase Completation of the extension process at the end of a chromosome Telomerase was found in abundance in cancer cells indicating that such enzyme is essential for cancer cells to continue dividing. Most cells in human undergo a limited number of cell divisions and then cease dividing. In these cells telomerase is basically not present. Telomerase binding proteins It is believed that after telomerase has extended the 3’-end a sufficient amount, DNA pol. a primes and DNA pol. d synthesizes a new Okazaki fragment.
Structure of origins of replication DnaA T antigen • The DNA sequences of replicators share two common features: • Binding sites (green) for initiation proteins that nucleates the assembly of replication machinery • A stretch of A-T rich DNA that unwinds readily but not spontaneously (blue) ORC Strategy for the identification of origins of replication in E. coli
In addition: * over-expression of DnaA leads to a burst of initiation. * under-expression of DnaA leads to increased initiation mass. v
E. coli replication origin oriC Mutants with a defect in any one of the genes for the main histone-like proteins (HU, FIS, H-NS, IHF) are viable. Even deficiency in two histone-like proteins is possible. Mutations in fis or him (IHF) genes result in asynchronous initiations. Cells depleted of HU, IHF and H-NS simultaneously are not viable. Model for initiation complex at oriC A compact complex is formed in the presence of HU protein and ATP which contains about 10 monomers of DnaA per oriC. The result of DnaA binding is that double helix melts within the 3 AT-rich, 13-nucleotides repeats. Base changes in individual DnaA boxes have surprisingly little effects. Any change of distances, however, inactivates oriC.
oriC The DnaA protein binds oriC A model for E. coli initiation of DNA replication The primase synthesizes the primer complementary to the leading strand Replication initiates bidirectionally. Initially only the leading strand is synthesized. After helicase has moved 1000 bases a second primer on each lagging strand is synthesized. Initiation of lagging strand synthesis.
For bacteria dividing more frequently than every 60 minutes, a cycle of replication must be initiated before the end of the preceding division cycle. At division (35/0 minutes), the cell receives a partially replicated chromosome. At 10 minutes, when this "old" replication fork has not yet reached the terminus, initiation occurs at both origins on the partially replicated chromosome. The start of these "new" replication forks creates a multiforked chromosome. Cells growing more rapidly are larger and possess a greater number of origins and there are correspondingly more chromosomes in the individual bacterium. Una cellula viene esposta per un breve tempo a [3H] timina (fase di pulse), poi viene somministrato un eccesso di timina fredda (fase di chase).
Replication of eukaryotic DNA Model for DNA replication DNA polymerases d and e are the main replicative enzymes
Switching delle Polimerasi Il prodotto del complesso primasi/DNA polimerasi è di circa 50-100 nucleotidi. Le DNA polimerasi d and ε sintetizzano rispettivamente il filamento guida e quello copia. The combined DNA polymerase and primase activities of the DNA polymerase a may constitute an evidence that this enzyme synthesizes the Okazaky fragments (??) but the DNA polymerase a has not the 3’ 5’ exonuclease activity. None of eukaryotic DNA pol. have the 5’ 3’ exonuclease activity like pol. I in E. coli.
Isolating the origins of yeast replicons (ARSs) S. cerevisiae mutants can be "transformed" to the wild phenotype by addition of DNA vectors that carries a wild-type copy of the gene (selection). Some yeast DNA fragments (when circularized) are able to transform defective cells very efficiently. These fragments can survive in the cell in the unintegrated (autonomous) state, that is, as self-replicating plasmids. A high-frequency transforming fragment possesses a sequence that confers the ability to replicate efficiently in yeast. This segment is called an ARS (autonomously replicating sequence).
Yeast origin of replication The subdomain B2 appears to correspond with the 13-nucleotide repeat array of E.coli. The subdomain B3 is the DNA binding site for ABF1 that by introducing a torsional stress, favours DNA melting at B2. 200 bp core region (40 bp) ORCs appear to remain attached to the yeast origins throughout the cell cycle (it is not the initiator proteins) and may have a key role in regulation of DNA replication. origin recognition complex ARS binding factor 1 Priming in eukaryotes In eukaryotes the Okazaki fragments are much shorter (200 bp) than in bacteria (about 4000 fragments of 1-2 Kb) and priming is a highly repetitive event. This indicates that each round of synthesis replicates the DNA associates with a single nucleosome (140-150 bp wound around the core particle plus 50-70 bp of linker). RPA (replication protein A) prevents duplex formation. 3’ 5’ DNA (50-100 bases) 10 bases RNA DNA pol. a DNA pol. d In eukaryotes the primase is an integral part of DNA pol. a. Possibly DNA synthesis (leading and lagging strands) involves the combined action of both DNA polymerasesd ande
Formation of pre-replicative complex Cdc6 multiple genome replication in absence of mitosis Cdc6 absence of pre-replication complexes Cdc6 protein was originally identified in yeast and subsequently shown to have homologs in higher eukaryotes Origin Recognition Complex ORC binds the origin Cdc6 is a highly unstable protein (half-life <5 minutes). Its rapid degradation means that no protein is available later in the cycle. The presence of Cdc6 in turn allows Mcm proteins to bind to the complex. Mutations in the Mcm factors could prevent initiation of replication. Helicase loading proteins Helicase Pre-Replicative Complex
Activation of the pre-replicative complex pre-RC is formed in G1. Kinases Cdk and Ddk are inactive in G1 and are activated when cells enter S phase. Phosphorylation of Cdc6 and Cdt1. Cdc6 and Cdt1 are displaced. Because these proteins are rapidly degraded during S phase, they are not available to support reloading of Mcm complex and so the origin cannot be used for a second cycle of initiation during the S phase. Synthesis of primer by Primase and positioning of the sliding clamp by PCNA protein. DNA Pol d recognizes primer and begins leading strand synthesis. DNA Pol d on the leading strand and DNA Pol e on the lagging strand.
Centinaia o addirittura migliaia di origini devono essere attivate una sola volta in ciascun ciclo. Le Cdk sono richieste per attivare il pre-RC. Una elevata attività Cdk promuove l’inizio della replicazione del DNA. L’attività delle Cdk inibisce la formazione di nuovi complessi pre-RC I livelli di Cdk variano durante il ciclo cellulare Gli elevati livelli di Cdk determinano la fosforilazione e conseguente dissociazione dall’origine di replicazione delle proteine Cdc6 e Cdt1 Gli elevati livelli di Cdk determinano l’associazione con l’origine di replicazione del solo ORC
Completation of lagging strand synthesis in eukaryotes None of the eukaryotic polymerase have a 5’ 3’ exonuclease activity. FEN1 endonuclease cannot initiate primer degradation because its activity is blocked by the triphosphate group present at the 5’-end of the primer. The flap model The RNase H model DNA-RNA junction