Download

1 / 33
460 likes | 1.1k Views
Nitric Oxide. NO refers to nitrosyl radical ( • NO) and its nitroxyl (NO – ) and nitrosonium (NO+) ions Freely diffusible, gaseous free radical. First described in 1979 as a potent relaxant of peripheral vasculature. Used by the body as a signaling molecule.

E N D
Nitric Oxide • NO refers to nitrosyl radical (•NO) and its nitroxyl (NO–) and nitrosonium (NO+) ions • Freely diffusible, gaseous free radical. • First described in 1979 as a potent relaxant of peripheral vasculature. • Used by the body as a signaling molecule. • Used as neurotransmitter, bactericide. • Environmental Pollutant • First gas known to act as a biological messenger N O
Nitric Oxide in plants • Affects aspects of plant growth and development. • Affects the responses to: light, gravity, oxidative stress, pathogens. • Can be a maturation and senescence factor • Has a concentration dependent cytotoxic or protective (antioxidant) effects.
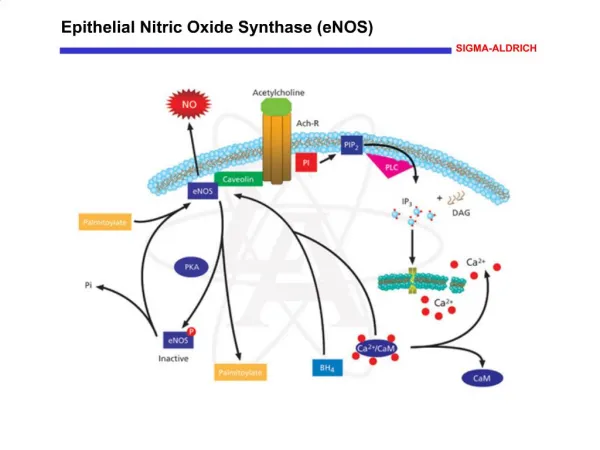
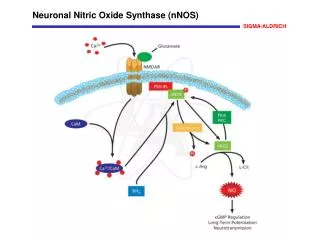
NO synthesis and reactions NO, a simple gas, is able to diffuse across the membrane, and alters the activity of intracellular target enzymes. The source of NO synthesis in plants includes reduction of nitrite by nitrate reductase (NR) and the oxidation of arginine to citrulline by NOS. No gene similar to mammalian NOS has been found. suggesting different NOS. AtNOS1 encodes a protein similar to snail NOS NO reversibly binds to ferrous iron within haemproteins, such as guanylate cyclase, haemoglobin and cytochrome oxidase. NO reacts at the diffusion limited rate with superoxide to produce peroxynitrite. NO or peroxynitrite, can react with thiols to produce nitrosothiols.
Chemistry & signaling of NO in biological environment NO signal transduction H2O2 and NO might regulate the activity of TFs directly via nitrosylation (NO) or oxidation of cysteine residues (H2O2). S-nitrosation–denitrosation is reversibleandcould represent a mechanism for regulating signal transduction. Abbreviations: M–NO, iron–nitrosyl complexes; M, heme- or iron-containing proteins; M–Fe(II)/Fe(III), Fe(II)/Fe(III)-containing metalloproteins; NOx, refers collectively to NO.2, N2O3 and N2O4; R–NO, nitrosated proteins Besides its half-life, a protein’s fate is manifest in its destination.
ROS – NO perturbations The Challenge - A molecule in a biological system encodes information in its shape, charge, hydrophobicity, and reactivity. Any change in a molecule’s composition encodes new information in the amount, rate, and duration of that change and where in the cell or organism the change takes place. • A large extent and diversity of NO-dependent modifications of biomolecules. Identification of these modifications has outraced our ability to gauge their physiological import. • STKE 2004 (2004) 52
Parallels between ROS and RNS in defense Feature ROS RNS Primary catalyst Multisubunit flavocytochrome Multisubunit flavocytochrome Substrates O2, NADPH O2, NADPH, L-arginine Primary product Inorganic radical (O2) Inorganic radical (·NO) Actions at low levels Activate or inhibit receptors, Activate or inhibit receptors, enzymes, enzymes, transcription transcription factors factors Actions at high levels Cause mutagenesis, apoptosis, necrosis Cause mutagenesis, apoptosis, necrosis Basis of celllar resistance SODs; catalase; peroxiredoxins; Under study redox cycles involving glutathione, thioredoxin, glutaredoxin, trypanothione,ovothione, mycothione; methionine sulfoxide reductase; ascorbate; -tocopherol; urate; -keto acids
2001 Nitric oxide-mediated signaling pathway in: NO inactivates IK,out. plant defense against pathogens Steady-state current-voltage curves derived from voltage clamp recordings (insets) from an intact guard cell before (○), during NO (■), and after washing (●). Curves are corrected for instantaneous current recorded at each voltage. Inset, Corresponding current traces cross-referenced by symbol with the voltage protocol
NO-induced cell death in Arabidopsis occurs independently of ROS • Cells were treated with methyl viologen (MV) to generate O2 · , NO donor (RBS), and/or the peroxynitrite scavenger and SOD-mimetic MnTBAP. • Cells were treated with RBS, H2O2 and/or catalase (CAT).
cGMP in NO-induced cell death The effects of the caspase-1 inhibitor Ac-YVAD-CMK on NO- and H2O2-induced cell death Cells were pre-treated with ODQ (guanylate cyclase inhibitor) and/or 8Br-cGMP prior to RBS.
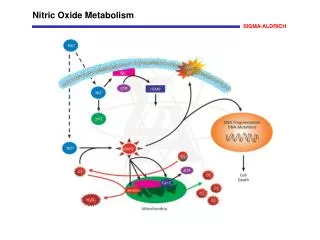
Main actions of nitric oxide (NO), peroxynitrite (ONOO−) and nitrosothiols (RSNO) on mitochondria. NO + Fe2+↔Fe2+−NO Reversible binding to haem a3. Responsible for inhibition at high ferrocytochrome c. concentrations−competitive with O2 NO + Cu2+ ↔ Cu+−NO+ Cu+−NO++H2O↔Cu++NO2−+2H+ Binding to Cu NO specifically and reversibly inhibits cytochrome oxidase (complex IV);nitrosothiols inactivate complex I; whereas peroxynitrite inhibits multiple respiratory complexes and aconitase, and activates the proton leak and permeability transition pore (ANT-PTP), which may contribute to NO-induced cell death. Inhibitions are indicated by dark arrows/bolts, while light ones indicate activations
The Role of O2 Work on the ryanodine receptor and mitochondrial respiration points to a reciprocal, concentration-dependent influence of O2 and NO on each other’s physiologic actions. In each case, NO exerts more control when the concentration of oxygen ([O2]) falls. A related concept was originally articulated for hemoglobin, where it was argued that S-nitrosylation controls O2 delivery, whereas [O2] controls S-nitrosylation.
Purification and Characterization of the Putative Tobacco NOS In vitro, a GST-AtNOS1 fusion protein was capable of NOS activity and showed similarities to mammalian NOS isoforms, but was not stimulated by typical mammalian NOS cofactors.
ABA- and nitrite-induced NO generation correlate with stomatal closure wild type (gray bars), abi1-1 (black bars), or abi2-1 (white bars) Stomatal apertures (black bars) wt (white bars)or NR mutant nia1, nia2 (black)
NO involvement in ABA-induced stomatal closure Effect of c-PTIO on ABA-induced stomatal closure
NO regulates K+ and Cl- channels in guard cells through a subset of ABA-evoked signaling pathways NO scavengers suppress ABA action in closing stomata NO donors promote it in the absence of ABA NO selectively inactivates IK+, in and Cl- out PNAS 2003 vol. 100 ; 11116 Voltage-clamp recordings from an intact guard cell (Inset) Steady-state I-V curves from voltage-clamp steps before (o), after 2 min of exposure to 10 µM SNAP (▲), and after washing in buffer-SNAP (●). ABA raises cytosolic-free [Ca2+] ([Ca2+]i) and cytosolic pH (pHi); these signals inactivate inward-rectifying K+ channels (IK,in) to prevent K+ uptake and activate outward-rectifying K+ channels (IK,out) and Cl- (anion) channels (ICl) at the plasma membrane to facilitate solute efflux +NO -NO NO promotes INTRACELLULAR Ca2+ releaseand thereby regulates guard cell ion channels
NO scavenger cPTIO blocks inactivation of IKin and activation of IClout by ABA and NO but not ABA-mediated activation of IKout Steady-state current for IK,out at +30 mV, IK,in at -200 mV, and ICl at -70 mV (shaded bars), effects of a 10-min exposure to ABA with or without 20 µM cPTIO (Left) and to 10 µM SNAP with or without 20 µM cPTIO (Right).
NOS activity and NO are reduced in Atnos1 mutants • NOS activity in leaf extracts of wild-type, Atnos1 mutant, and rescued Atnos1 plants. L-NAME inhibits the NOS activity in wild-type and rescued Atnos1 plants. • AtNOS1 mRNA levels in roots after treatment with ABA for 30 min in dark do not increase substantially. • C to H) NO production (shown as green fluorescence from the NO-sensitive dye DAF-2 DA) is increased by ABA in roots of wild-type seedlings and is reduced in Atnos1 mutants with or without 50 µM ABA. L-NAME (200 µM) inhibits ABA-induced NO production Science 302 (2003) 100 - 3.
AtNOS1 functions in ABA-induced stomatal closure (A) ABA induces NO production in guard cells of wt and rescued Atnos1 plants but not Atnos1 mutants, as shown by the increase in fluorescence from the NO-sensitive dye DAF-2 DA. L-NAME inhibits the ABA-induced NO production in all three genotypes. (B) Rel fluorescence signal from guard cells corresponding to treatments in (A) (n = 8) are shown. (C) ABA-induced stomatal closure is inhibited in Atnos1 mutants and is partially restored in rescued Atnos1. (D) ABA fails to inhibit light-induced stomatal opening in Atnos1 mutants but functions well in rescued Atnos1 mutant plants.
The mutant is impaired in leaf greening, shoot growth, and fertility Phenotypes of the Atnos1 mutant and complementation with a 35S-AtNOS1. (A) Wt and (B) Atnos1 mutants with yellow first true leaves (C) Atnos1 containing a 35S-AtNOS1 transgene (rescued Atnos1) has green true leaves. (D) Atnos1 mutation inhibits shoot growth (E) Root development is inhibited in Atnos1 mutants grown on agarose. (F) Reproductive growth and fertility are reduced in Atnos1 mutants. (G) Western blots of leaf extracts show that the Atnos1 mutation reduces AtNOS1 protein levels. Equal amount of protein loaded (AtNRT1.1 control). (H to K) Treatment of Atnos1 mutants with NO restored greening of first true leaves, whereas treatment with sodium ferrocyanide (a SNP analog that does not produce NO) had no effect
Results- cell viability GA-induced PCD Exogenous NO donor delays PCD ABA-inhibits PCD
NO delays the GA-induced loss of CAT and SOD(mRNA+protein) Plant Physiol, August 2002, Vol. 129, pp. 1642-1650
NO and Cell Death +PBITU Psm (avrRpm 1) NO and H2O2 cause cell death NO and O2- react to form peroxynitrite % Cell Death Peroxynitrite (ONOO -) does not cause cell death Too much O2- ‘mops up NO’ – no death Delladonne et al. (2001) PNAS 98:13454
BHT delays GA-induced PCD in barley aleurone layers Aleurone layers were incubated with GA alone or with GA plus 1 mM BHT, and were then loaded with the fluorescent probes FDA and FM 4-64 at 24, 36, and 48 h
SUMMARY • NO delays the onset of GA-induced PCD of barley aleurone cells. • NO may act to protect the cell from oxidative stress as an antioxidant. • NO has cytotoxic and protective effects in plants. • NO delays the GA-induced loss of CAT and SOD and can be an endogenous modulator of aleurone cell viability. • NO is produced in barley aleurone cells. • But, we can not specify the exact mechanism by which NO exerts its protective effect.
Arabidopsis flowering time September 26, 2004 • Two Arabidopsis plants, sown at the same time. The left is already flowering, whereas on the right it is delayed because it carries active FRI. • An electron micrograph of an extremely early flowering transgenic plant overexpressing the floral pathway integrators FT and LFY. • The consequence of loss of floral meristem identity function. The lfy mutant (left) has leaf-like structures in place of the flowers ( right)
Multiple Pathways Control Flowering TimeIntegration of the Multiple Inputs Depending on the wavelength of light, the light quality the inputs promote or repress activation of genes termed floral pathway integrators. FRI repression prevents promotive photoperiod (PP) accelerating flowering in late summer or autumn. Vernalization (VRN) antagonizes FRI repression, reducing FLC activity thus enabling long days in spring to upregulate the floral path integrators (FPI) Smaller circles and thinner arrows indicate a lesser role Science, Vol 296, 285-289 , 12 April 2002
Air pollutant nitric oxide acts as a plant hormone to delay flowering in plants. The scientists discovered that while plants produce their own internal nitric oxide to regulate flowering, they are also influenced by external concentrations of the chemical.
Exogenous NO promotes vegetative growth but inhibits reproductive development The effects of an NO donor SNP on plant growth and development. Arabidopsis seedlings were grown in SNP during long days (16h light/8h dark) for 5 weeks. (B) [SNP] on growth (C and D) The effect of SNP on flowering times. Fresh weight per shoot (B), the rosette leaf number (C), and days to bolting (D) from experiments as in (A) and fig. S1A plotted as a function of the concentrations of SNP that were applied, respectively Science, Vol 305, 1968-1971 , 24 September 2004
Involvement of cGMP in the flowering regulation • The involvement of cGMP in the regulation of the flowering of Pharbitis was investigated by exogenous applications of cGMP and chemicals that change the cGMP level and analyses of endogenous cGMP level. • There was a significant difference in the cyclic GMP level between 16-h-long night conditions and a long night with a night-break. During a long inductive night the oscillation of cGMP was observed with four main peaks in 4, 7, 11, 14 h, whereas a 10 min flash of red light in the middle of the night was able to modify these rhythmical changes in the second half of the long night. These results have shown that there are oscillations in the concentration of cGMP in the night and the biosynthesis and/or deactivation of cGMP is affected by light treatment and therefore it may be involved in the regulation of photoinduction processes in cotyledons. • From these combined results, we propose a hypothesis that cGMP is involved in the control of photoperiodic flower induction in Pharbitis nil. (J Plant Physiol, 2004, March vol. 161. 277-284)
Endogenous NO represses the floral transition NO overproducer (nox) A) The root growth phenotype in nox1 mutant. B) Endogenous NO levels in nox1 and WT. White-light images are shown at the bottom. C) The levels of L-Arg and NO in WT, nox1, and cue1-5. Plants grown under 12h light/12h dark cycles were harvested 6h after dawn. D) The nox1 mutant flowers late. WT and nox1 were grown in soil under 12-hour light/12-hour dark cycles and were photographed after 60 days of growth. (E and F) Flowering times of nox1 and cue1-5 mutants. The rosette leaf number (E) and days to flowering (F) from experiments as in (D) were scored. G) The NO synthase 1 (nos1) mutant that produces fewer NO flowers early under long days. (H) The nos1 mutant flowers earlier than WT under SNP treatments.
NO affects the expression of genes that control the floral transition. NO suppresses the photoperiod pathway