Download
1 / 1
10 likes | 196 Views
Post-Translationally Modified Cardiac Mitochondrial VDAC1 Gating Kinetics Analyzed Using Continuous-Time MCMC Model. Shivendra G. Tewari a , Yifan Zhou b , Amadou K.S. Camara c , Ranjan K. Dash a,d , and Wai-Meng Kwok b,c

E N D
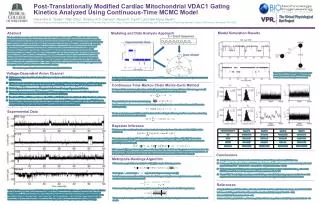
Post-Translationally Modified Cardiac Mitochondrial VDAC1 Gating Kinetics Analyzed Using Continuous-Time MCMC Model Shivendra G. Tewaria, Yifan Zhoub, Amadou K.S. Camarac, Ranjan K. Dasha,d, and Wai-Meng Kwokb,c aBiotechnology and Bioengineering Center, bDepartment of Pharmacology and Toxicology,cDepartment of Anesthesiology, and dDepartment of Physiology, Medical College of Wisconsin, Milwaukee, WI 53226 Model Simulation Results Modeling and Data Analysis Approach Abstract Mitochondrial homeostasis, maintained via the outer mitochondrial membrane (OMM), is critical for maintaining mitochondrial function. The voltage-dependent anion channel (VDAC1 isoform) is the most abundant protein on the OMM, and it regulates the exchange of ions and molecules across the membrane. VDAC1 possesses a remarkable ability to conduct metabolites and anions in its high conductance state and cations in its low conductance state. Despite its significance, no integrated mitochondrial bioenergetics model incorporates for VDAC1. Our electrophysiological recordings of VDAC1 incorporated into lipid bilayers revealed that post-translational modification (PTM) by phosphorylation and nitration significantly impacted VDAC1 gating kinetics. Consequently, a thorough statistical treatment was applied by modeling VDAC1 activity using a generalized continuous-time Markov chain Monte Carlo (MCMC) method. The MCMC method returns probability distribution of identified model parameters from which important model information such as appropriateness and parameterization can be easily obtained. Our analyses suggest that phosphorylation and nitration of VDAC1 facilitate multiple conductance states wherein a simple kinetic model incorporating three open states (O1 ↔ O2 ↔ O3) describes the experimental data. The long-term goal is to link the changes in VDAC1 gating kinetics to mitochondrial function. The work is supported by NIH/R01-HL095122 and NIH/P50-GM094503. Voltage-Dependent Anion Channel Three-state WT VDAC model. Rate constants for phosphorylated and nitrated VDAC recordings were also estimated using the same model. • Highly conserved protein of the mitochondrial outer membrane which facilitates transport of solutes of molecular mass below 5000 Da. • VDAC is the main conduit that connects the cytosol to the mitochondrial inter-membrane space, and allows the passages of substrates (e.g. pyruvate, succinate), phosphates (e.g. ATP, ADP, Pi), and cations (e.g. Ca2+, K+, H+) across the OMM. • VDAC1 oligomerization concurrent with apoptosis induction and is stopped by apoptosis blockers. • Recent studies have shown that VDAC undergoes PTM in neurodegenerative diseases such as Huntington’s and Alzheimer’s disease, and also in cardiac ischemia-reperfusion (IR) injury. • The effect of PTMs on VDAC are not known. How PTMs on VDAC affect mitochondrial and cellular function is not known. • Herein, we developed a continuous-time Markov Chain Monte Carlo (MCMC) model for single channel VDAC1 kinetics. The MCMC model qualitatively describes the effect of PTMs and nitration on VDAC gating kinetics. This will enable us to computationally study the effect of mutant VDAC on mitochondrial and cellular function. A schematic diagram communicating the basic concept of MCMC parameter estimation from the single channel current recordings of reconstituted VDAC1 protein. Continuous-Time Markov Chain Monte-Carlo Method For the model shown in above schematic, a matrix Q is generated whose entry equal the transition rates qij or zero (if the states are not connected). Following conservation, the diagonal entries are chosen to be: The probability of each state is calculated using: To define transitions from one state to another during a sampling interval, , we need transition probability matrix which is given by: We also need stationary probabilities which is obtained after solving the following set of linear equations and making sure the detailed balance condition holds: Experimental Data Bayesian Inference Whether a random model explains the random data better is given by the conditional probability P(Q|E) which is evaluated as: M is used as an auxiliary variable that enables us to generate model samples (Q) from P(Q, M|E). The transition probability from a Mk to Mk+1 are available from the transition probability matrix A therefore the above mentioned probability is computed as [2]: Siekmann et al. [2] proposed a variant of forward-backward algorithm which considers just two conductance states of a channel. Their algorithm was modified to account for different conductance states of VDAC seen in our experiments. It is done using different projection matrices, for different conductance states of VDAC, while realizing M. Conclusions Metropolis-Hastings Algorithm • A simple three state mechanism is found suitable to explain wild-type and mutant VDAC kinetics. • Residue-specific phosphorylation and ONOO- induced nitration, both, lead to an increased conductivity of the half-open VDAC state: WT – 1.4 nS, T70E – 1.6 nS, S137E – 1.9 nS, and Nitration – 2.0 nS. • T70E phosphorylated VDAC remains more closed than WT and other mutants evident from it’s q21 value while S137E has a much faster activity compare to it’s rate constants with the WT. • Nitrated VDAC has the highest conductance in it’s full-open state (2.8 nS) and stays open most of the time which might contribute to the increased oxidative stress, as previously suggested by [4] in Alzheimer’s disease. MH is used to sample different model Q from P(M, Q|E) using the following equation: From a given , a new sample is generated using a two-step process [3]: In the first step a new sample model is generated and after generating the new sample it is made sure that the condition of detailed balance is satisfied: In the second step, the new generated sample is accepted or rejected based on the following criterion: The new sample is accepted with 100% probability if it’s likelihood of generating observed events is more than the old sample. References 1. Yang M, AKS Camara, BT Wakim, Y Zhou, AK Gadicherla, W-M Kwok, DF Stowe (2012), Biochim Biophys Acta 1817: 2049-2059. 2. Siekmann I, LE Wagner, D Yule, C Fox, D Bryant, EJ Crampin, J Sneyd (2011), Biophys J 100: 1919 – 1929. 3. Brooks SP (1998), The Statistician 47: 69 – 100. 4. Sultana R, HF Poon, J Cai, WM Pierce, M Merchant, JB Klein, WR Markesbery, DA Butterfield (2006), Neurobiol Dis 22: 76 – 87. VDAC-1 cDNA was over-expressed in E. Coli and purified using Ni-NTA HisBind Resins and refolded by drop-wise dilution plus dialysis at 4 C. Channel activity of purified VDAC-1 protein was measured by reconstituting it into planar lipid bilayer. VDAC currents were measured at a voltage clamp of ±10 mV. Phospho-mimetic mutants S137E and T70E were constructed by substituting serine 137 and threonine 70 with glutamate which mimic phospho-serine and phospho-threonine. Nitration of reconstituted VDAC was induced by 50 mol/L of exogenous peroxynitrite (ONOO-) [1].