Download

1 / 35
370 likes | 674 Views
The Organization and Expression of Ig Genes. Justin Walter Immunology, Fall 2008. Introduction . The vertebrate immune system is capable of responding to an essentially infinite array of foreign antigens Variable vs. constant regions

E N D
The Organization and Expression of Ig Genes Justin Walter Immunology, Fall 2008
Introduction • The vertebrate immune system is capable of responding to an essentially infinite array of foreign antigens • Variable vs. constant regions • Basis for variability organization and expression of Ig genes
Overview • Historical perspective – early theories • “Multigene” organization of Ig genes • Variable region gene rearrangements • Basics of mechanism – key players: DNA signal sequences, specific enzymes • Generation of Ab diversity • seven primary routes
Historical perspective • Ig sequence analysis revealed many dilemmas • Extreme diversity of Ab specificity • Variable regions vs constant regions • Isotypes with similar Ag specificity but differing heavy-chain constant regions
Historical perspective • Proponents of the one-gene-one-protein paradigm had trouble reconciling this model with the oddities of Igs. • This led to an initial “germ-line theory” which suggested that a significant portion of the genome is dedicated solely to Ab coding. • Argument: the immune system is THAT important
Historical perspective • In contrast, “somatic-variation theories” emerged which suggested the opposite: • Relatively small amount of Ig genes • Specificity arises from mutation and/or recombination
Historical perspective • Dreyer & Bennet (1965) • Two-gene, one-protein model • No precedent in any biological system
Historical perspective • Tonegawa & Hozumi (1976) • Compare Ig DNA from embryonic (germline) and adult myeloma (somatic) cells • Experimental data suggested that during differentiation, the V and C genes undergo rearrangement. • 1987 Nobel prize
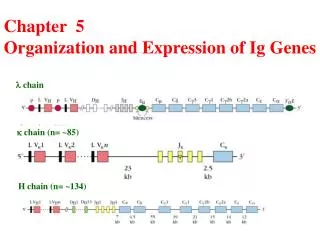
Multigene Organization of Ig genes • Each “class” of Ig components (kappa, lambda, heavy) encoded by separate multigene families on different chromosomes • Each family contains several coding sequences, or gene segments
Multigene Organization of Ig genes • Κ & λlight chains: V (variable), J (joining), and C (constant) gene segments • Heavy chains: V, D (diversity), J, and C gene segments • A leader (L) sequence also precedes each V segment. • Gene segments discovered by comparing DNA sequences with amino acid sequences of Igs • Tonegawa, again.
Organization of Ig germ-line gene segments (mouse) Pre-rearrangement!
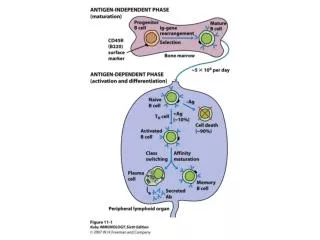
Variable-region rearrangement • Multifaceted process, produces mature B cells which are “committed” to express specific Ab • Specificity of Ab determined by the sequence of its rearranged variable genes.
Light-chain rearrangements • V-J rearrangements • Specific “allowed” rearrangements differ from species to species, but a “big-picture” view can suffice • Rearrangement occurs in ordered steps but can be considered as random events which result in the random determination of Ab specificity
Kappa light-chain rearrangement & RNA processing Leader sequence targets nascent protein to ER and is subsequently cleaved
Heavy-chain rearrangements • Requires two separate rearrangement events • D-J joining • V-DJ joining • Differential polyadenylation & RNA splicing can result in mRNA with either Cu or Cδ • heavy chain genes • B cells can express BOTH IgM and IgD with identical Ag specificity on its surface
Heavy-chain rearrangement The focus here is on B cells… IgM IgD
Mechanism of Variable region DNA rearrangements • Recombination signal sequences direct recombination
Mechanism of Variable region DNA rearrangements • Recombination signal sequences direct recombination Vλ, JK, VH, JH VK, Jλ, DH
Mechanism of Variable region DNA rearrangements • A one-turn RSS can only join with a two-turn RSS • Why might this be?
Mechanism of Variable region DNA rearrangements • Gene segments are joined by a class of enzymes called recombinases • Two recombination-activating genes encode proteins which act together to mediate V-(D)-J joining • RAG-1 • RAG-2
Same transcriptional orientation • (most common) b. opposite transcriptional orientation Addn. of P-nucleotides accomplished with repair enzymes
Mechanism of Variable region DNA rearrangements • Rearrangements may be productive or nonproductive.
Mechanism of Variable region DNA rearrangements • Allelic exlusion • Heavy-chain genes only expressed from one chromosome • Light-chain genes only expressed from one chromosome • Essential for specificity • Expression of both alleles would result in a “multispecific” B cell
Mechanism of Variable region DNA rearrangements • Allelic exlusion
Generation of Ab diversity • Multiple germ-line gene segments • Combinatorial V-(D)-J joining • Junctional flexibility • P-region nucleotide addition • N-region nucleotide addition • Somatic hypermutation • Combinatorial association of light and heavy chains
Somatic hypermutation • Nucleotide replacement, mediated by activation-induced cytidine deaminase (AID) • Also plays a key role in class switching • Frequency of 10-3 per bp per generation • 100,000X the rate of spontaneous mutation! • Approx 1 mutation every 2 cell divisions