Download

1 / 75
820 likes | 1.17k Views
Chapter 19 The Tricarboxylic Acid Cycle. Outline. What is the chemical logic of the TCA cycle ? How is pyruvate oxidatively decarboxylated to Acetyl-CoA ? How are two CO 2 molecules produced from Acetyl-CoA ? How is oxaloacetate regenerated to complete the TCA cycle ?

E N D
Outline • What is the chemical logic of the TCA cycle ? • How is pyruvate oxidatively decarboxylated to Acetyl-CoA ? • How are two CO2 molecules produced from Acetyl-CoA ? • How is oxaloacetate regenerated to complete the TCA cycle ? • What are the energetic consequences of the TCA cycle ?
Outline Can the TCA cycle provide intermediates for biosynthesis ? What are the anaplerotic, or “filling up,” reactions ? How is the TCA cycle regulated ? Can any organisms use acetate as their sole carbon source ?
Oxidation of glucose to CO2 is a 24-electron oxidation The electrons from glucose oxidation feed into the electron transport pathway, driving synthesis of ATP.
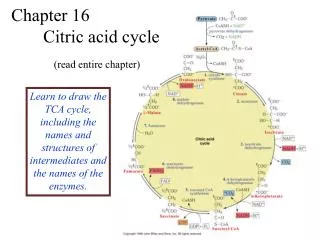
Hans Krebs Showed That the Oxidation of Acetate is Accomplished by a Cycle The TCA cycle is known as the Krebs cycle – and also as the citric acid cycle • Pyruvate from glycolysis is oxidatively decarboxylated to acetate. • Then acetate is degraded to CO2 in the TCA cycle. • Some ATP is produced. • More NADH is made. • NADH goes on to make more ATP in electron transport and oxidative phosphorylation.
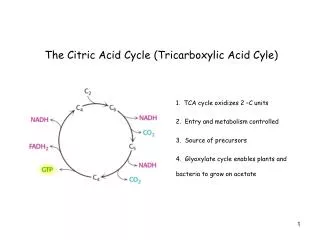
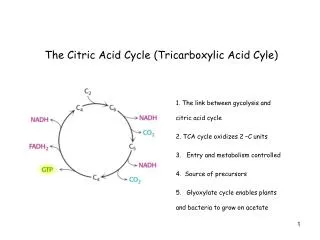
19.1 What Is the Chemical Logic of the TCA Cycle? • The TCA cycle seems like a complicated way to oxidize acetate units to CO2. • What are the possibilities (based on our knowledge of metabolism) ? • cleavage between Cs and to a carbonyl. • an-cleavage of an -hydroxyketone. • These methods cleave C-C bonds and oxidize but do not work for acetyl-CoA.
Cleavage Between Carbon Atoms and to a Carbonyl • This type of cleavage occurs in the fructose bisphosphate aldolase reaction in glycolysis. • But it will not work with acetate, which does not have a β carbon.
-Cleavage of an -Hydroxyketone • This type of cleavage occurs in the transketolase reaction (described in Chapter 22). • But it will not work with acetate, which does not have an α-hydroxyl group.
The Chemical Logic of TCA – a better way to cleave acetate • Living things have evolved the clever chemistry of condensing acetate with oxaloacetate and then carrying out a β-cleavage. • TCA combines this with oxidation to form CO2, regenerating oxaloacetate and capturing all the energy as NADH and ATP ! • Pyruvate must enter the mitochondria to enter the TCA cycle. • Oxidative decarboxylation of pyruvate is catalyzed by the pyruvate dehydrogenase complex. • Pyruvate dehydrogenase is a noncovalent assembly of three enzymes. • Five coenzymes are required.
E.Coli Pyruvate Dehydrogenase (PDH) Ecoli PDH is a cubic complex, 4 x 106 daltons. The center of the cube (core) contains 24 chains of E2 (dihydrolipoyl transacetylase). E2 requires lipoamide and Coenzyme A. Each edge of the cube (12) contains a dimer of E1 (pyruvate dehydrogenase). E1 requires thiamine pyrophospate (TPP•Mg++). Each face of the cube (6) contains a dimer of E3 (dihydrolipoyl dehydrogenase). E3 requires FAD and NAD+.
Liver PDH is a Multi-Subunit Complex of Three Enzymes, 9 x 106 daltons Figure 19.3 Icosahedral model of the pyruvate dehydrogenase complex (PDC) core structure. E1 subunits are yellow; E2 subunits are green. Linkers in blue; E3 not shown.
Figure 19.4 The Reaction Mechanism of the Pyruvate Dehydrogenase Complex Decarboxylation of pyruvate yields hydroxyethyl-TPP. Transfer to lipoic acid is followed by formation of acetyl-CoA.
The Coenzymes of the Pyruvate Dehydrogenase Complex Thiamine pyrophosphate. TPP assists in the decarboxylation of α-keto acids (here) and in the formation and cleavage of α-hydroxy ketones (as in the transketolase reaction; see Chapter 22).
The Coenzymes of the Pyruvate Dehydrogenase Complex The Nicotinamide Coenzymes – NAD+/NADH and NADP+/NADPH carry out hydride (H:-) transfer reactions. All reactions involving these coenzymes are two-electron transfers.
The Coenzymes of the Pyruvate Dehydrogenase Complex The Flavin Coenzymes – FAD/FADH2 & FMN/FMNH2. Flavin coenzymes can exist in any of three oxidation states, and this allows flavin coenzymes to participate in one-electron and two-electron transfer reactions. Partly because of this, flavoproteins catalyze many reactions in biological systems and work with many electron donors and acceptors.
The Coenzymes of the Pyruvate Dehydrogenase Complex • Coenzyme A. • The two main functions of Co A are: • Activation of acyl groups for transfer by nucleophilic attack. • Activation of the α-hydrogen of the acyl group for abstraction as a proton. The reactive sulfhydryl group on CoA mediates both of these functions. The sulfhydryl group forms thioester linkages with acyl groups. The two main functions of CoA are illustrated in the citrate synthase reaction (see Figure 19.6).
The Coenzymes of the Pyruvate Dehydrogenase Complex Lipoic Acid functions to couple acyl-group transfer and electron transfer during oxidation and decarboxylation of α-keto acids. It is found in the pyruvate dehydrogenase and α-ketoglutarate dehydrogenase complexes. Lipoic acid is covalently bound to relevant enzymes through amide bond formation with the ε-NH2 group of a lysine side chain.
Figure 19.5 The mechanism of the first three steps of the pyruvate dehydrogenase complex reaction
CAC Reactions Events and in the Citric Acid Cycle • C-C bond formation -31.4 kJ/mol • Isomerization +6.7 kJ/mol • NAD+ dependent decarboxylation -8.4 kJ/mol • NAD+ dependent decarboxylation -30.0 kJ/mol • GTP formation -3.3 kJ/mol • FAD linked dehydrogenation +0.4 kJ/mol • Hydration -3.8 kJ/mol • NAD+ dependent dehydrogenation +29.7 kJ/mol
19.3 How Are Two CO2 Molecules Produced from Acetyl-CoA? The citrate synthase reaction initiates the TCA cycle Citrate synthase shows classic CoA chemistry. Cα of the acetyl group in acetyl-CoA is acidic and can be deprotonated to form a carbanion. This carbanion is a strong nucleophile that can attack the α-carbonyl of oxaloacetate, yielding citryl-CoA. Thioester hydrolysis then produces citrate. NADH & succinyl-CoA are allosteric inhibitors. Note (Table 19.1) that citrate synthase has a large negative G and is a site of regulation.
The citrate synthase synthase reaction initiates the TCA cycle Figure 19.6 Citrate is formed in the citrate synthase reaction from oxaloacetate and acetyl-CoA. The mechanism involves nucleophilic attack by the carbanion of acetyl-CoA on the carbonyl carbon of oxaloacetate, followed by thioester hydrolysis.
The citrate synthase synthase reaction initiates the TCA cycle Four of these are oxidation reactions, two of which are oxidative decarboxylations. The other four include a C-C bond synthesis, an isomerization, a substrate level phosphorylation and a hydration.
The citrate synthase synthase reaction initiates the TCA cycle Figure 19.7 Citrate synthase in mammals is a dimer of 49-kD subunits. In the monomer shown here, citrate (blue) and CoA (red) bind to the active site, which lies in a cleft between two domains and is surrounded mainly by α-helical segments.
Citrate Is Isomerized by Aconitase to Form Isocitrate • Citrate is a poor substrate for oxidation. • So aconitase isomerizes citrate to yield isocitrate which has a secondary-OH, which can be oxidized. • Note the stereochemistry of the reaction: aconitase removes the pro-R H of the pro-R arm of citrate. • Aconitase uses an iron-sulfur cluster - see Figure 19.8.
Citrate Is Isomerized by Aconitase to Form Isocitrate Figure 19.8 The aconitase reaction converts citrate to cis-aconitate and then to isocitrate. Aconitase is stereospecific and removes the pro-R hydrogen from the pro-R arm of citrate.
Citrate Is Isomerized by Aconitase to Form Isocitrate Figure 19.8 The active site of aconitase. The iron-sulfur cluster (pink) is coordinated by cysteines (orange) and isocitrate (purple).
Fluoroacetate Blocks the TCA Cycle • Fluoroacetate is an extremely poisonous agent that blocks the TCA cycle in vivo, although it has no apparent effect on any of the isolated TCA cycle enzymes. • The action of fluorocitrate has been traced to aconitase • Aconitase is inhibited by fluorocitrate, which is formed from fluoracetate in two steps, as shown here.
Aconitase Utilizes an Iron-Sulfur Cluster Figure 19.9 The iron-sulfur cluster of aconitase. Binding of Fe2+ to the vacant position of the cluster activates aconitase. The added iron atom coordinates the C-3 carboxyl and hydroxyl groups of citrate and acts as a Lewis acid, accepting an electron pair from the hydroxyl group and making it a better leaving group.
Aconitase Utilizes an Iron-Sulfur Cluster Other iron-sulfur cluster types:
Isocitrate Dehydrogenase Catalyzes the First Oxidative Decarboxylation in the Cycle Oxidative decarboxylation of isocitrate yields -ketoglutarate • This is classic NAD+ chemistry (hydride removal) followed by a decarboxylation. • Isocitrate dehydrogenase is a link to the electron transport pathway because it makes NADH. • The mechanism is typical of NAD+-dependent enzymes (see Figure 19.10). • The reaction involves (first) oxidation of the C-2 alcohol of isocitrate to form oxalosuccinate, then a β-decarboxylation reaction that expels the central carboxyl group as CO2.
Isocitrate Dehydrogenase Catalyzes the First Oxidative Decarboxylation in the Cycle Figure 19.10 The isocitrate dehydrogenase reaction. Hydride removal by NAD+ is followed by a β-decarboxylation reaction that expels the central carboxyl group as CO2. Similar in mechanism to glucose-6-P dehydrogenase of the hexose shunt which requires NADP+.
The Subunits of α-Ketoglutarate Dehydrogenase are Similar to Those of Pyruvate Dehydrogenase This multienzyme complex is very similar in assembly and function to the pyruvate dehydrogenase complex.
-Ketoglutarate Dehydrogenase Catalyzes the Second Oxidative Decarboxylation of the TCA Cycle A second oxidative decarboxylation • This enzyme is a self-assembling complex nearly identical (structurally and mechanistically) to pyruvate dehydrogenase. • Five coenzymes used - TPP•Mg++, CoASH, Lipoic acid, NAD+, FAD. • This is the least reversible reaction of the cycle and is energetically irreversible.
-Ketoglutarate Dehydrogenase Catalyzes the Second Oxidative Decarboxylation of the TCA Cycle Like pyruvate dehydrogenase, -ketoglutarate dehydrogenase is a multienzyme complex – consisting of -ketoglutarate dehydrogenase, dihydrolipoyl transsuccinylase, and dihydrolipoyl dehydrogenase. The complex uses the same five different coenzymes.
Conversion of SuccinoylCoA to Succinate Succinyl-CoA Synthetase Catalyzes a Substrate-Level Phosphorylation • A nucleoside triphosphate is made. • This is possible because succinyl-CoA is a high-energy thioester intermediate. • Its hydrolysis (the hydrolysis of a CoA ester) drives the phosphorylation of GDP to produce GTP. • The mechanism (Figure 19.11) involves a phosphohistidine.
Conversion of SuccinoylCoA to Succinate Hydrolysis of succinyl-CoA (a CoA ester) drives the phosphorylation of GDP to produce GTP. Like glycolysis, the CAC has one reaction that requires inorganic phosphate, Pi.
Mechanism • Figure 19.11 The mechanism of the succinyl-CoA synthetase reaction. • Phosphate anion carries out a nucleophilic attack on the carbonyl C of succinyl-CoA, forming succinyl phosphate • Subsequent phosphoryl transfer to an active-site His forms a phosphohistidine intermediate, releasing the product succinate • The phosphoryl group is then transferred to GDP to produce GTP
Conversions seen in other Pathways • Conversions seen in the next three reactions will be seen five times throughout the text. • These reactions include: • Oxidation of a single bond to a double bond. • Hydration across the double bond. • Oxidation of the resulting alcohol to a ketone. • This trio of reactions will be seen again in: • Fatty acid breakdown and synthesis. • Amino acid breakdown and synthesis.
Succinate Dehydrogenase Is an FAD-Dependent Oxidation An FAD-dependent oxidation of a single bond to a double bond • The mechanism involves hydride removal by FAD and a deprotonation. • This enzyme is actually part of the electron transport pathway and is bound to the inner mitochondrial membrane. • The electrons transferred from succinate to FAD (to form FADH2) are passed to ubiquinone (UQ) in the electron transport pathway via iron-sulfur clusters.
The Succinate Dehydrogenase Reaction Since succinate is symmetrical any label initially in acetylCoA will be scrambled between the ends of the remaining metabolites.
Succinate Dehydrogenase Uses Iron-Sulfur Clusters Succinate Dehydrogenase contains three types of Fe-S centers – a 4Fe-4S center, a 3Fe-4S center, and a 2Fe-2S center, as shown here.
FAD is Covalently Bound to Succinate Dehydrogenase Figure 19.12 FAD is covalently bound to succinate dehydrogenase. The C-8a carbon of FAD is linked to the N-3 of a His residue of the enzyme.
Fumarase Catalyzes the Trans-Hydration of Fumarate to Form L-Malate Hydration occurs across the newly formed double bond catalyzed by fumarase • Hydration involves trans-addition of the elements of water across the double bond. • The enzyme is stereospecific. It produces only L-malate from the trans double bond. • Possible mechanisms are shown in Figure 19.13. • The actual mechanism is not known for certain.
Possible Mechanisms For the Fumarase Reaction Figure 19.13
Malate Dehydrogenase Completes the Cycle by Oxidizing Malate to Oxaloacetate An NAD+-dependent oxidation • The carbon that gets oxidized is the one that received the-OH in the previous reaction. • This reaction is energetically expensive Go' of +30 kJ/mol. • Thus the concentration of oxaloacetate in the mitochondrial matrix is quite low. • However, the malate dehydrogenase reaction is pulled forward by the favorable citrate synthase reaction.

















![Tricarboxylic acid cycle (TCA Cycle) [Kreb’s cycle] [Citric acid cycle]](https://cdn3.slideserve.com/6696193/tricarboxylic-acid-cycle-tca-cycle-kreb-s-cycle-citric-acid-cycle-dt.jpg)
