Download

1 / 13
130 likes | 136 Views
Although lidocaine is a commonly used local anesthetic in dental treatment, the effects of lidocaine on calcium release in craniofacial arteries such as the lingual artery are not well known.

E N D
Journal Home Page www.bbbulletin.org BRITISH BIOMEDICAL BULLETIN Original The Effects of Lidocaine on Calcium Release and the Role of Pathways in Swine Lingual Artery Contraction Induced with Agonists Kenichi Satoh*, Mami Chikuda and Shigeharu Joh Division of Dental Anesthesiology, Department of Reconstructive Oral and Maxillofacial Surgery, School of Dentistry, Iwate Medical University, Japan A R T I C L E I N F O Received 30 June 2015 Received in revised form 05 July 2015 Accepted 12 July 2015 Keywords: Lidocaine, Caffeine, Intracellular Ca2+ concentration, Inositol trisphosphate channel, L-type channel. Corresponding author: Division of Dental Anesthesiology, Department of Reconstructive Oral and Maxillofacial Surgery, School of Dentistry, Iwate Medical University, Japan. E-mail address: satoken@iwate- med.ac.jp A B S T R A C T Background: Although lidocaine is a commonly used local anesthetic in dental treatment, the effects of lidocaine on calcium release in craniofacial arteries such as the lingual artery are not well known. Aims: The aim of this study was to examine the effects of lidocaine on calcium release and the role of pathways in swine lingual artery contraction induced with agonists. Materials and Methods: We measured intracellular Ca2+ concentration ([Ca2+]i)and tension using front-surface fluorometry in sections of swine lingual artery with denuded endothelium. Results: The [Ca2+]i and tension induced with adrenaline and histamine in the absence of extracellular Ca2+ with lidocaine added were low compared with or without lidocaine, while the [Ca2+]i and tension induced with caffeine were the same with or without lidocaine. Treatment with lidocaine before and during the application of adrenaline significantly inhibited the increase in [Ca2+]i and tension induced with adrenalinein the presence of extracellular Ca2+ after depletion of the intracellular Ca2+ store. Conclusions: Lidocaine depressed increases in [Ca2+]i and tension that were dependent on the Ca2+ via inositol trisphosphate channel- operated Ca2+-entry channels, and lidocaine did not attenuate Ca2+- induced Ca2+ release in KCl- and agonist-induced smooth muscle contraction. Lidocaine depressed the increase of Ca2+ influx from extracellular Ca2+ through RACC or nonselective cation channels.
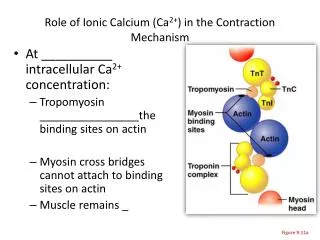
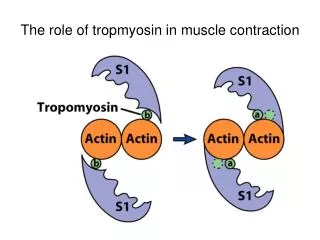
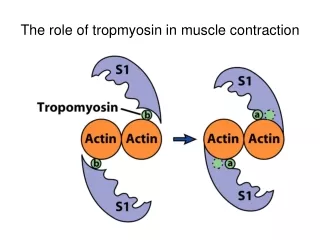
Satohet al______________________________________________________ ISSN-2347-5447 Introduction Lidocaine is widely used local anesthetic agents; it is administered in various surgical operations, in the treatment of ventricular arrhythmia, suppression of airway reflexes associated with tracheal intubation. Despite common usage of lidocaine by dentists as a local anesthetic in dental treatment and in oral and maxillofacial surgery, reports1,2 on the effect of lidocaine on craniofacial arteries. Among craniofacial arteries, the lingual artery is relatively unexplored though it is a prominent artery. “Characterization of the contraction of lingual artery must reveal its features both common to and distinct from those of other vascular beds, which may have implications for the clinical treatment for pathological circulation in the tongue”1. Therefore, it is important for dentists to understand the mechanism of vasodilation induced by lidocaine in lingual artery. “Lidocaine vascular muscle, airway smooth muscle and other smooth muscles, with the mechanism of the direct relaxant effect of lidocaine on smooth muscle potentially caused by an effect on Ca2+ mobilization”2,3. However, the contraction of vascular smooth muscle is regulated by changes (intracellular) Ca2+ levels ([Ca2+]i) and Ca2+ sensitivity of contractile “Depolarization of the sarcolemma with a high concentration of KCl causes the influx of extracellular Ca2+ through voltage-gated Ca2+ channels, whereas binding of agonists such as noradrenaline to the receptors causes the release of Ca2+ from intracellular Ca2+ stores such as the inositol trisphosphate (IP3) channel-operated store or one of the sarcoplasmic stores”5,6. Agonists release Ca2+ from intracellular Ca2+ stores to induce an initial transient contraction (phasic type), followed by activation of Ca2+ influx to induce sustained contraction (tonic type)7. Aims of this present study were to investigate the effects of lidocaine on calcium release and the role of pathways in this process in swine lingual artery contraction induced by agonists. Material and Method and for This study was approved by the Institutional Review Committee on the Ethics of Animal Experiments of Iwate Medical University (approval number 26- 010). All experiments were conducted in accordance with the Institutional Animals Care and Use Committee guidelines. Reagents and solutions L-adrenaline purchased from Tokyo Chemical Industry Co., Ltd. (Tokyo, Japan). All other chemicals were obtained from Wako Pure Chemical Industries (Osaka, Japan). In all experiments, air-equilibrated Hanks’ balanced salt solution (HBSS) was used to keep arteries at resting conditions. HBSS consisted of 137 mM NaCl, 5.4 mM KCl, 0.8 mM MgSO4, 1.26 mM CaCl2, 0.34 mM Na2HPO4, 0.44 mM KH2PO4, 4.2 mM NaHCO3, and 5.55 mM glucose (pH 7.34). All other salt solutions (SSs) used as perfusate were made by modifying HBSS. SS containing 100 mM was made by substituting the respective concentration of KCl for the equivalent concentration of NaCl in HBSS. Ca2+-free SS was made by adding GEDTA (to give a concentration of 1 mM) and omitting CaCl2 from HBSS. Each SS containing adrenaline, ryanodine, and verapamil alone or in combination was made by adding the component agents into the respective base SS or Ca2+-free SS. Artery ring preparation and contraction measurement In swine tongues, segments of lingual artery were prepared and isometric there are few bitartrate was generally relaxes in cytosolic elements4. caffeine, BBB[3][3][2015] 304-316
Satohet al______________________________________________________ ISSN-2347-5447 tension was measured as described in detail previously1,2. Fresh swine tongues were obtained from a local abattoir. A segment of lingual artery was cut from the proximal region of the tongue. After adventitia was removed, artery segments about 2 mm in diameter were cut into 2-mm-long rings, and the lumen surface was rubbed gently against the thin arm of stainless steel tweezers to remove endothelium. An artery ring was held with two tungsten needles in a perfusion chamber with a holding volume of perfusate of 2 ml. One needle was fastened to a displacement transducer (Type UL-2GR, Minebea Co., Fujisawa, Japan), and the other to a micromanipulator. The solution was bubbled with a mixture of 95 % O2 and 5 % CO2, and held at a temperature of 37 °C and flow rate of 1.6 ml/min with a peristaltic pump (SMP- 23, Tokyo Rikakikai Co., Tokyo, Japan). As the strength of contraction did not change when the resting tone was 3–7 mN, the artery rings were extended to give a resting tone of approximately 5 mN. After a 30-min HBSS perfusion, artery rings were perfused with 60 mM KCl for 2 min as a control. After a 10–20-min HBSS perfusion to obtain a resting position, artery rings were perfused with HBSS or Ca2+-free SS plus a stimulant for 5 min either alone or in combination with lidocaine. Contractions increases in isometric tension with the displacement transducer, detected were amplified with a carrier amplifier (CSD-815 Minebea Co., Fujisawa, Japan) and recorded with a Powerlab 16/30T data acquisition system (AD Instruments, Australia). The contraction strengths in each experiment were normalized to the strength of the 60 mM KCl contraction and expressed as a percentage. Fura-2 loading Artery rings were loaded with the Ca2+ indicator dye, fura-2, by incubation in HBSS containing 10-µM fura-2/AM (an acetoxymethyl ester form of fura-2) for 3 h at 37 °C in the dark. After loading with fura- 2, the rings were washed with HBSS to remove the dye in the extracellular space, and were then kept in HBSS for at least 1 h before the start of measurements. Measurement of fura-2 fluorescence Changes in fluorescence intensity of the fura-2-Ca2+ complex were monitored using a front-surface fura-2 fluorometer (Aquacosmos, Hamamatsu Photonics K.K., Tokyo, Japan). Two wavelengths of excitation light (340 and 380 nm) were obtained spectroscopically from a Xenon light source. Rings were illuminated by guiding the two alternating wavelengths of excitation light through quartz optic fibers. Surface fluorescence of the ring was collected by glass optic fibers and introduced through a 500-nm emission light, which was induced by alternating two wavelengths of excitation light (340 nm and 380 nm). The ratio of the fluorescence intensity (fluorescence ratio) at 340-nm excitation (F340) to that at 380-nm excitation (F380) was monitored to estimate changes in [Ca2+]i; these changes were expressed as a percentage, with the values at rest in HBSS (5.84 mM K+) and 60 mM KCl designated as 0 % and 100 %, respectively. Simultaneous measurement of tension and fluorescence ratio A final 2-min 60 mM KCl perfusion was done after each experiment to confirm that each artery ring had retained intact contractility throughout the experiment. The strength of any contractions and change in [Ca2+]i in an experiment was normalized to the strength of 60 mM KCl contraction and were detected as and signals Digital indicator, BBB[3][3][2015] 304-316
Satohet al______________________________________________________ ISSN-2347-5447 fluorescence ratio, and expressed as a percentage. Effect of lidocaine on the contraction induced with 60 mM KCl or 5 µM adrenaline in artery rings with denuded endothelium. Artery rings in either 60 mM KCl or 5 µM adrenaline were perfused with lidocaine concentrations of 0.1 µM, 1.0 µM, 10 µM, 100 µM, 0.5 mM, 1.0 mM, 5.0 mM and 10 mM for 2 min. Effect of lidocaine on the contraction induced with adrenaline in the absence of extracellular Ca2+ in artery rings with denuded endothelium. 5 µM adrenaline in Ca2+-free SS was perfused for 5 min. Effect of lidocaine on the contraction induced with histamine in the absence of extracellular Ca2+ in artery rings with denuded endothelium. 5 µM histamine in Ca2+-free SS was perfused for 5 min. Effect of lidocaine on the contraction induced with caffeine in the absence of extracellular Ca2+ in artery rings with denuded endothelium. 25 mM caffeine in Ca2+-free SS was perfused for 5 min. Effect of lidocaine on the contraction induced with adrenaline in the presence of extracellular Ca2+ after depletion of the intracellular Ca2+ store in artery rings with denuded endothelium. Artery rings were placed in Ca2+-free SS containing 1 mM EDTA for 5 min. To remove Ca2+ from the intracellular Ca2+ stores, 25 mM caffeine was applied three times in the same Ca2+-free SS. To achieve this, 25 mM “first caffeine” was applied for 5 min in Ca2+-free SS, followed by a 5 min washout with Ca2+-free SS. Then 6 mM ryanodine was applied with Ca2+-free SS for 5 min, followed by the “second caffeine” application for 5 min. After another washout with Ca2+-free SS for 5 min, the “third caffeine” was applied for 5 min followed by washout for 5 min using Ca2+-free SS. Subsequently, adrenaline in Ca2+-free SS was applied for 15 min in HBSS. Statistical analysis Values are presented as mean ± SEM. Differences between the means of two groups were evaluated for significance using the Student’s t test. Differences between two among multiple groups, whose homogeneity of variance was assessed by the Levene test, were evaluated using Dunnet’s t test. Differences were considered significant at P < .05. Results Effect of lidocaine on the contraction induced with 60 mM KCl or 5 µM adrenaline in artery rings with denuded endothelium Lidocaine attenuated the contraction and [Ca2+]i induced by 60 mM KCl or 5 µM adrenaline in a concentration-dependent manner (Fig. 1). Effect of lidocaine on the contraction induced with adrenaline in the absence of extracellular Ca2+ in artery rings with denuded endothelium Figure 2 shows a representative time course of [Ca2+]i and tension development induced by applications of 60 mM KCl and 5 µM adrenaline in Ca2+-free SS. The application of 5 µM adrenaline in Ca2+-free SS for 5 min in the absence of extracellular Ca2+ caused a transient increase in [Ca2+]i and tension. The increases in [Ca2+]i and tension that occurred with the addition of lidocaine were low compared with those that occurred without lidocaine addition. The respective peak increases in [Ca2+]i with and without lidocaine were 14 ± 2 % and 35 ± 2 % (n = 8), and the respective peak increases in tension with and without lidocaine were 37 ± 2 % and 76 ± 1 % (n = 8) (Table 1). BBB[3][3][2015] 304-316
Satohet al______________________________________________________ ISSN-2347-5447 Effect of lidocaine on the contraction induced with histamine in the absence of extracellular Ca 2+ in artery rings with denuded endothelium Figure 3 shows a representative time course of [Ca2+]i and tension development induced by applications of 60 mM KCl and 5 µM histamine in Ca2+-free SS. The application of 5 µM histamine for 5 min in the absence of extracellular Ca2+ caused a transient increase in [Ca2+]i and tension in Ca2+-free SS. The increases in [Ca2+]i and tension that occurred with the addition of lidocaine were low compared with those that occurred without lidocaine addition. The respective peak increases in [Ca2+]i with and without lidocaine were 21 ± 3 % and 71 ± 2 % (n = 8), and the respective peak increases in tension with and without lidocaine were 16 ± 1 % and 69 ± 2 % (n = 8) (Table 1). Effect of lidocaine on the contraction induced with caffeine in the absence of extracellular Ca2+ in artery rings with denuded endothelium Figure 4 shows a representative time course of [Ca2+]i and tension induced by applications of 60 mM KCl and 25 mM caffeine in Ca2+-free SS. The application of 5 µM caffeine for 5 min in the absence of extracellular Ca2+ caused a transient increase in [Ca2+]i and tension in Ca2+-free SS. The increases in [Ca2+]i and tension that occurred with the addition of lidocaine were the same as those that occurred without lidocaine. The respective peak increases in [Ca2+]i with and without lidocaine were 42 ± 3 % and 42 ± 2 % (n = 8), and the respective peak increases in tension with and without lidocaine were 40 ± 2 % and 42 ± 2 % (n = 8) (Table 1). Effect of lidocaine on the contraction induced with adrenaline in the presence of extracellular Ca2+ after depletion of the intracellular Ca2+ store in artery rings with denuded endothelium Figure 5 shows a representative time course of [Ca2+]i and tension development induced by applications of 60 mM KCl and 5 µM adrenaline in HBSS after depletion of the intracellular Ca2+ store. The application of 5 µM adrenaline for 15 min in the presence of extracellular Ca2+ caused [Ca2+]i and tension to develop slowly to peak strength, with respective peak increases in [Ca2+]i and tension of 39 ± 2 % (n = 8) and 83 ± 2 % (n = 8). Treatment with 1 mM lidocaine 5 min before and during the application of adrenaline inhibited increases in [Ca2+]i and tension induced by the application of adrenaline; peak increases in [Ca2+]i and tension were 23 ± 3 % (n = 8) and 37 ± 2 % (n = 8), respectively (Fig. 5 B and Table 1). Discussion significantly In the present study we found out three important issues. Lidocaine depressed the increase in [Ca2+]i and the tension induced with KCl and adrenaline in a concentration-dependent manner. Lidocaine depressed the increase in [Ca2+]i and tension induced with adrenaline and histamine, but did not depress the increase in [Ca2+]i and tension induced by caffeine in the absence of extracellular Ca2+. Lidocaine also depressed the increase in [Ca2+]i and tension induced with adrenaline in the presence of extracellular Ca2+ after depletion of the intracellular Ca2+ store. First, lidocaine increase in [Ca2+]i and the tension induced with KCl and adrenaline in a concentration- dependent manner. The contraction of vascular smooth muscle is controlled primarily by sarcoplasmic concentration. When sarcoplasmic Ca2+ depressed the Ca2+ BBB[3][3][2015] 304-316
Satohet al______________________________________________________ ISSN-2347-5447 levels rise, calmodulin forms a complex with Ca2+ that activates myosin light-chain (MLC) kinase, resulting phosphorylation, which leads to force generation1,8. The increase in [Ca2+]i occurs via the influx of extracellular Ca2+ and the release of Ca2+ from sarcoplasmic stores9. It has been recognized that smooth muscle contraction is due to Ca2+ influx through voltage-dependent and channels9,10. It is generally assumed that “KCl-induced smooth muscle contraction depends largely on Ca2+ influx through voltage-gated Ca2+ channels, and agonist- induced smooth muscle contraction depends on Ca2+ release stores”1,8.” Ca2+-induced (CICR) also contributes to Ca2+ release from intracellular Ca2+ stores during KCl- and agonist-induced smooth muscle contraction, and receptor-activated (RACC) or receptor-regulated nonselective cation channels that conduct Na+, K+ and Ca2+ are involved in the Ca2+ influx from extracellular Ca2+ in agonist-induced smooth muscle contraction”11,12. Based on this knowledge, we suggest that lidocaine may depress the increase of Ca2+ influx through voltage-gated Ca2+ channels and CICR in KCl-induced smooth muscle contraction, and inhibit Ca2+ release from sarcoplasmic stores and RACC in agonist-induced smooth muscle contraction. Second, lidocaine depressed the increase in [Ca2+]i and tension induced with adrenaline and histamine but did not depress the increase in [Ca2+]i and tension induced by caffeine in the absence of extracellular Ca2+. To understand vascular contraction induced with agonists, we must consider factors including the vascular bed, stimulant, and contractile type classified as phasic (fast) or tonic (slow). Contraction is reported to consist of two components: the phasic contraction appears as a large increase in the initial part of contraction, and the tonic contraction appears as a sustained level of force in the lag phase1. Contraction induced with agonists depends on Ca2+ release from sarcoplasmic stores8. It is generally assumed that the phasic contraction depends on Ca2+ release from sarcoplasmic stores8, and the tonic contraction depends largely on Ca2+ influx from extracellular Ca2+ via RACC9,13. In the absence of extracellular Ca2+, we observed whether lidocaine depressed the increase in [Ca2+]i and tension that were dependent on Ca2+ sarcoplasmic stores. “noradrenaline- and contraction depended on Ca2+ via IP3 channel-operated Ca2+ channel on the sarcolemma”1,5,6. We suggest that lidocaine depressed the increases in [Ca2+]i and tension that were dependent on the Ca2+ via IP3 channel-operated Ca2+ channels and CICR. Caffeine releases Ca2+ from the intracellular Ca2+ stores and increases the sensitivity of CICR to Ca2+10,14. However, lidocaine did not depress the increase in [Ca2+]i and tension induced by caffeine in the absence of extracellular Ca2+. We suggest that lidocaine did not attenuate CICR in KCl- and agonist-induced smooth muscle contraction. In other words, in KCl- induced contraction, lidocaine may mainly depress the increase of Ca2+ influx through voltage-gated Ca2+ channels. From the above observation, adrenaline, histamine and caffeine can release Ca2+ from the same Ca2+ intracellular store since this store can be depleted by caffeine. However, two previous studies reported that the caffeine- and the noradrenaline-sensitive Ca2+ stores were different from each other because the rate of depletion and recovery of these stores were different14,15. Furthermore, noradrenaline-sensitive Ca2+ store is too small to produce contraction directly; Ca2+ from the noradrenaline-sensitive store may release Ca2+ from the caffeine-sensitive Ca2+ in MLC receptor-linked release In histamine-induced from general, from sarcoplasmic Ca2+ release Ca2+ channels we suggest that the BBB[3][3][2015] 304-316
Satohet al______________________________________________________ ISSN-2347-5447 store by the CICR mechanism and thus lead to contraction. It is still unclear whether the intracellular Ca2+ store compartment. Third, lidocaine increase in [Ca2+]i induced with adrenaline in the presence of extracellular Ca2+ after depletion of the intracellular Ca2+ store. Tonic contraction depends on Ca2+ influx through RACC or nonselective cation channels9-11. In this study, it was suggested that lidocaine depressed the increase of Ca2+ influx from extracellular Ca2+ through RACC or nonselective cation channels. KCl and noradrenaline appear to open the same L-type Ca2+ channels9,11. Therefore, it was possibility that lidocaine depressed the increase in Ca2+ influx from extracellular Ca2+ via L-type channels. In conclusion, lidocaine depressed the increase in [Ca2+]i and tension induced with KCl and adrenaline in a concentration- dependent manner, and depressed the increase in these induced with adrenaline and histamine. In contrast, lidocaine did not depress the increase in [Ca2+]i and tension induced by caffeine in the absence of extracellular Ca2+. However, lidocaine did depress the increase in [Ca2+]i and tension induced with adrenaline in the presence of extracellular Ca2+ after depletion of the intracellular Ca2+ store. Therefore, it was suggested that lidocaine depressed the increase in Ca2+ via IP3 channel-operated Ca2+ channels and CICR, that lidocaine did not attenuate CICR in KCl- and agonist- induced smooth muscle contraction, and that lidocaine depressed the increase of Ca2+ influx from extracellular Ca2+ through RACC or nonselective cation channels. Further investigation is needed into whether the intracellular Ca2+ store is a single compartment in the lingual artery. Acknowledgements Conflict of interest The all authors declare that they have no conflicts of interests. Abbreviations [Ca2+]i: Intracellular Ca2+ concentration IP3 channel-operated trisphosphate channel-operated store NA: Noradrenaline HBSS: Hanks’ balanced salt solution SS: Salt solution Fura-2/AM: Acetoxymethyl ester form of fura-2 CICR: Ca2+-induced Ca2+ release RACC: Receptor-activated Ca2+ channels. References 1. Kawaguchi T, Satoh K, Kuji A, Joh S. Features of distinct contractions induced with a high and a low concentration of KCl, noradrenaline, and histamine in swine lingual artery. Archives of Pharmacology. 2013; 381: 107– 120. doi:10.1007/s00210-009-0486-9. 2. Satoh K, Kamada S, Kumagai M, Sato M, Kuji A and Joh S. Effect of lidocaine on swine lingual and pulmonary arteries. J of Anesth. 2015. [Epub ahead of print] 3. Kai T, Nishimura J, Kobayashi S, Takahashi S, Yoshitake J, Kanaide H. Effects of lidocaine on intracellular Ca2+ and tension in airway smooth muscle. Anesthesiology. 1993 78: 954–965. 4. Karaki H. Cytosolic calcium levels in vascular smooth muscle. Nihon Ketsueki Gakkai Zasshi. 1989; 52: 1506–1515. 5. Janiak R, Wilson SM., Montague S, Hume JR. Heterogeneity of calcium stores and elementary release pulmonary arterial smooth muscle cells. Am J Physiol Cell Physiol. 2011; 280: 22–33. 6. Ureña J, Smani T, López-Barneo J. Differential functional properties of Ca2+ stores in pulmonary arterial conduit and resistance myocytes. Cell Calcium. 2004; 36: 525–534. 7. Karaki H, Sato K, Ozaki H. Different effects of verapamil on cytosolic Ca2+ and is a single depressed the store: Inositol Naunyn-Schmiedeberg's events in canine None. BBB[3][3][2015] 304-316
Satohet al______________________________________________________ ISSN-2347-5447 12.Sato K, Ozaki H, Karaki H. Multiple effects of caffeine on contraction and cytosolic free Ca2+ levels in vascular smooth muscle of rat aorta. Naunyn Pharmacol. 1988; 338: 443–448. 13.Ozaki H, Ohyama T, Sato K, Karaki H. Ca2(+)-dependent mechanisms of sustained contraction in vascular smooth muscle of rat aorta. Jpn J Pharmacol. 1990; 52: 509-512. 14.Mori T, Yanagisawa T, Taira N. Histamine increases vascular tone and intracellular calcium level using both intracellular and extracellular calcium in porcine coronary arteries. Jpn J Pharmacol. 1990; 52:263-271. 15.Saida K, van Breemen C. Characteristics of the norepinephrine-sensitive Ca2+ store in vascular smooth muscle. Blood Vessels. 1984; 21: 43-52. contraction in norepinephrine-stimulated vascular smooth muscle. Jpn J Pharmaco. 1991; 55: 35–42. Ogut O, Brozovich FV. Regulation of force in vascular smooth muscle. J Mol Cell Cardiol. 2003; 35: 347–355. Karaki H, Ozaki H, Hori M, Mitsui-Saito M, Amano K, Harada K, et al. Calcium movements, distribution, and functions in smooth muscle. Pharmacol Rev. 1997; 49: 157–230. 10.Karaki H, Sato K, Ozaki H. Different effects of norepinephrine and KCl on the cytosolic Ca2+-tension relationship in vascular smooth muscle of rat aorta. Eur J Pharmacol. 1988; 151: 325–328. 11.Ozaki H, Karaki H. Different Ca (2+)- sensitivity in phasic and tonic types of smooth muscles. Biol Signals. 1993; 2:253– 262. 8. Schmiedebergs Arch 9. and independent Table 1. The effect of lidocaine on swine lingual artery ring contraction induced with agonists in the presence or absence of extracellular Ca2+ Adrenaline without lidocaine [Ca2+]i (%) Adrenaline in the absence of extracellular Ca2+ Histamine in the absence of extracellular Ca2+ Caffeine in the absence of extracellular Ca2+ Adrenaline in the presence of extracellular Ca2+ after depletion of intracellular Ca2+ Values represent the mean ± SEM. [Ca2+]i: intracellular concentration of Ca2+. * denotes a significant difference compared with the value obtained without lidocaine addition. Adrenaline without lidocaine Tension (%) Adrenaline with lidocaine [Ca2+]i (%) Adrenaline with lidocaine Tension (%) 35 ± 2 76 ± 1 14 ± 2* 37 ± 2* 71 ± 2 69 ± 2 21 ± 3* 16 ± 1* 42 ± 2 42 ± 2 42 ± 3 40 ± 2 39 ± 2 83 ± 2 23 ± 3* 37 ± 2* BBB[3][3][2015] 304-316
Satohet al______________________________________________________ ISSN-2347-5447 Figure 1. The effect of a variety of lidocaine concentrations on swine lingual artery ring contraction induced by a) 60 mM KCl or b) 5 µM adrenaline Lidocaine attenuated the contraction and [Ca2+]i induced by 60 mM KCl or 5 µM adrenaline in a concentration-dependent manner (n=8). The fluorescence ratio was monitored to estimate changes in intracellular Ca2+ concentration. BBB[3][3][2015] 304-316
Satohet al______________________________________________________ ISSN-2347-5447 Figure 2. The effect of lidocaine on swine lingual artery contraction induced with adrenaline in the absence of extracellular Ca2+ This is a representative time course of fluorescence ratio (an estimate of [Ca2+]i) and tension development induced by applications of 60 mM KCl and 5 µM adrenaline in Ca2+-free salt solution. The application of 5 µM adrenaline for 5 min in the absence of extracellular Ca2+ caused a transient increase in [Ca2+]i and tension in Ca2+-free SS salt solution. BBB[3][3][2015] 304-316
Satohet al______________________________________________________ ISSN-2347-5447 Figure 3. The effect of lidocaine on swine lingual artery ring contraction induced by histamine in the absence of extracellular Ca2+ This is a representative time course of fluorescence ratio (an estimate of [Ca2+]i) and tension development induced by applications of 60 mM KCl and 5 µM histamine in Ca2+-free salt solution. The application of 5 µM histamine for 5 min in the absence of extracellular Ca2+ caused a transient increase in [Ca2+]i and tension in Ca2+ free salt solution. The increases in [Ca2+]i and tension that occurred with the addition of lidocaine were low compared with those that occurred without lidocaine addition. BBB[3][3][2015] 304-316
Satohet al______________________________________________________ ISSN-2347-5447 Figure 4. The effect of lidocaine on swine lingual artery ring contraction induced by caffeine in the absence of extracellular Ca2+ This is a representative time course of fluorescence ratio (an estimate of [Ca2+]i) and tension development induced by applications of 60 mM KCl and 25 mM caffeine in Ca2+-free salt solution. The application of 5 µM caffeine for 5 min in the absence of extracellular Ca2+ caused a transient increase in [Ca2+]i and tension in Ca2+-free salt solution. The increases in [Ca2+]i and tension that occurred with the addition of lidocaine were the same as those that occurred without lidocaine addition. BBB[3][3][2015] 304-316
Satohet al______________________________________________________ ISSN-2347-5447 Figure 5. The effect of lidocaine on swine lingual artery ring contraction induced by adrenaline in the presence of extracellular Ca2+ after depletion of intracellular Ca2+ stores This is a representative time course of fluorescence ratio (an estimate of [Ca2+]i) and tension development induced by applications of 60 mM KCl and 5 µM adrenaline in Hanks’ balanced salt solution after depletion of the intracellular Ca2+ store. The application of 5 µM adrenaline for 15 min in the presence of extracellular Ca2+ caused [Ca2+]i and tension to develop slowly to peak strength. Treatment with 1 mM lidocaine 5 min before and during the application of adrenaline significantly inhibited the increases in [Ca2+]i and tension induced by the application of adrenaline. BBB[3][3][2015] 304-316