Download
1 / 62
630 likes | 934 Views
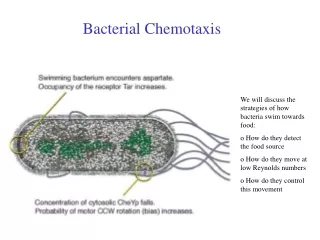
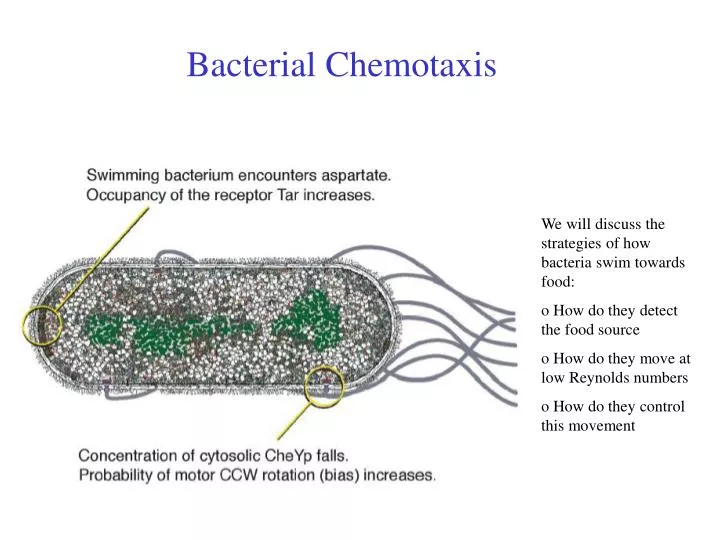
Bacterial Chemotaxis. We will discuss the strategies of how bacteria swim towards food: o How do they detect the food source o How do they move at low Reynolds numbers o How do they control this movement. Chemotaxis. Definition :

E N D
Bacterial Chemotaxis We will discuss the strategies of how bacteria swim towards food: o How do they detect the food source o How do they move at low Reynolds numbers o How do they control this movement
Chemotaxis Definition : „Die gerichtete Bewegung von Organismen hin oder weg von chemischen Lock- bzw. Schreckstoffen.“ Chemotaxis bei Bakterien Salmonella typhimurium werdenvon Serin angelockt (links) und von Phenol abgeschreckt (rechts) Die Amöbe Dictyostelium Discoideum läuft auf steigende Konzentrationen von cAMP zu
External Polymers in Bacteria Typically, two different External Polymers are found in Bacteria: o Bacteria can move with Pili by extending and retracting them inside the cell body (Type IV pilia). They are used to glue bacteria to each other to form biofilms, connect different bacteria for exchanging plasmids or let them attach to surfaces. o Flagella are used to swim. Often, Bacteria have several Flagella.
Flagella Motor Bacteria swim with the help of Flagella (Geißel) which are rotated by transmembrane motors. A Flagella can rotate with up to 150 turns per second.
Flagella Motor Movie http://video.google.com/videoplay?docid=-6630027279608718766
Bacteria move by rotating the Flagella At small scales, viscosity of water dominates and effects by inertia are absent. Thus swimming strategies are very much different from what we know at large scales. A good strategy is to swim by "drilling holes with a screw".
Moving and Tumbling Bacteria Depending on the Rotation direction, several Flagella either form a bundle and allow forward simming or they "repell" each other and block swimming. The latter is due to inward propagation of interaction defects, given by the geometry of the flagella.
Moving and Tumbling Bacteria From: Real-Time Imaging of Fluorescent Flagellar Filaments, LINDA TURNER, WILLIAM S. RYU, AND HOWARD C. BERG, JOURNAL OF BACTERIOLOGY, May 2000, p. 2793–2801
Strategie der Chemotaxis • E. coli Bakterien haben effiziente molekulare Motoren zur Fortbewegung • Sie bewegen sich durch abwechselnde Schwimm- und Taumel Bewegung
Bakterielle Chemotaxis Die bakteriellen Geißeln sind in Bündeln angeordnet, welche die Eigenschaft haben, dass sie sich im Uhrzeigersinn (CW) drehen auseinanderstreben und sich zusammenlagern, wenn sie sich gegen den Uhrzeigersinn (CCW) drehen. Das Bakterium kennt daher zwei Bewegungszustände (A) geradeaus schwimmen (v=14-30m/sec, im Mittel 0.8s) und (B) taumeln. Im Mittel für 0.2s. Wird für das Bakterium die Konzentration eines Lockstoffes höher mit der Zeit, wird das Taumeln unterdrückt. Dadurch bewegt sich das Bakterium im Mittel in Richtung zunehmender Konzentrationen.
Schwimm/Taumel (run & tumble) -Bewegung Die Bewegung von E.coli ist eine Folge von geradliniger Fortbewegung gefolgt von kurzen Umorientierungsphasen. Diese Schwimm&Taumel-Bewegung kann als Diffusionsprozess mit exponentiel verteilter Schrittlänge vt beschrieben werden. Man erhält die effektive Diffusionskonstante: Laufgeschwindigkeit v, mittlerer Laufzeit Richtungsfaktor a. Richtungsfaktor a beschreibt die mittlere Neuorientierung nach jeder Taumelphase =<cos>. Eine Gleichverteilung aller Richtungen (random walk) entspricht =0, keine Richtungsänderung =1. Mit v=30m/sec und = 0,8sec, =0 erhält man DST=240 µm2/sec. Im Vergleich kleine Bio-Moleküle (Zucker, Peptide etc.) : D=400 µm2/sec !!!
Mechanismus der mittleren chemotaktischen Drift Eine Erhöhung der Lockstoffkonzentration führt zu einer vorübergehenden Verlängerung der „Schwimmphase“. Dadurch wird der random walk ungleich gewichtet und es ergibt sich einemittlere Driftgeschwindigkeit in Richtung höherer Lockstoffkonzentration. => Biased Random Walk Constraints: - only~ 1,000receptors - only1 sto evaluate concentration Performance: - can detect < 1%change of occupancy - can operate over 5 orders of magnitude in ambient concentration
Optical Methods: Optical Tweezer Ecoli in water.avi
Tethering on Substrates Ecoli tethered.avi
System-Eigenschaften des biochemischen Netzwerks • Messung einer Konzentrationsänderung, nicht einer absoluten Konzentration (Zeitliche Messungen) • Exakte Anpassung • Empfindlichkeit durch Verstärkung • Signalkombination unterschiedlicher Attraktoren und Schreckstoffe Der Konzentrationsbereich, welcher Chemotaxis auslöst erstreckt sich über 5 Größenordnungen (nM->mM) !
Input : Attraktorkonzentration Output :Taumelbewegung
Das intrazelluläre Signal-Netzwerk Aus genetischen Untersuchungen sind vier cytoplasmatische Proteine CheA, CheW, CheY und CheZ bekannt, die die chemotaktischen Rezeptoren mit dem Geißelmotor verbinden. CheW: Hilfsfaktor CheA : Proteinkinase CheY : Response Regulator CheZ : Phosphatase-activating Protein Reguliert durch den Rezeptor transferiert CheA eine Phosphorylierung auf das kleine CheY Protein. CheY-p diffundiert zu den Motorproteinen der Flagellen und induziert den Übergang des Motors in die Taumelbewegung. (CheY-p = Ein/Ausschalter)
Das intrazelluläre Signal-Netzwerk Phosphorylation is the addition of a phosphate (PO4) group to a protein molecule or a small molecule. Reversible phosphorylation of proteins is an important regulatory mechanism. Enzymes called kinases (phosphorylation) and phosphatases (dephosphorylation) are involved in this process. Many enzymes are switched "on" or "off" by phosphorylation and dephosphorylation. Reversible phosphorylation results in a conformational change in the structure in many enzymes. For example, phosphorylation replaces neutral hydroxyl groups on serines, threonines or tyrosines with negatively charged phosphates with pKs near 1.2 and 6.5. Thus, near pH 6.5 they add 1.5 negative charges and above pH 7.5 they add 2 negative charges. It is estimated that 1/10th to 1/2 of proteins are phosphorylated (in some cellular state). A phosphorylated serine residue (from: http://en.wikipedia.org/wiki/Phosphorylation
Das intrazelluläre Signal-Netzwerk CheR : Methyltransferase CheB : Methylesterase Die Rezeptoren werden mittels Methylierung sensitiver. Die Methylierung macht das Protein CheR, die desensitisierende Demethylierung das phosphorylierte CheB. Die Phosphorylierung von CheB wird von CheA getriggert. Damit ergibt sich an negativer Feedback-Loop. Werden Rezeptoren detektiert, wird über CheA und p-CheB der Rezeptor weniger sensitiv. Methylation denotes the attachment or substitution of a methyl group on various substrates. Besides DNA methylation, typically at CpG sequence sites, Protein methylation typically takes place on arginine or lysine amino acid residues. Arginine can be methylated once or twice, Lysine can be methylated once, twice or three times. Protein methylation is one type of post-translational modification. From: http://en.wikipedia.org/wiki/Methylation
Das intrazelluläre Signal-Netzwerk Phosphorylation is the addition of a phosphate (PO4) group to a protein molecule or a small molecule. Reversible phosphorylation of proteins is an important regulatory mechanism. Enzymes called kinases (phosphorylation) and phosphatases (dephosphorylation) are involved in this process. Many enzymes are switched "on" or "off" by phosphorylation and dephosphorylation. Reversible phosphorylation results in a conformational change in the structure in many enzymes.
Bakterien adaptieren ihre Signalantwort Beobachtung nach Zugabe des Lockstoffs bei t=0 : Die Taumelfrequenz (Häufigkeit einer Taumelbewegung pro Sek.) nimmt schlagartig ab. Nach einer gewissen Adaptionszeit tsteigt die Frequenz wieder auf den alten Wertes an. Vorteil: Bakterium kann Veränderung der Lockstoffkonz. über einen enormen Größenbereich 10-10 M -10-3 M detektieren. unstimulated cells Systemantwort ist schnell <200ms, Adaption langsam (10 Minuten) cells stimulated at time t =0
Adaption durch Methylierung Methyl-akzeptierende Rezeptoren (MCP) Es gibt 8 Methylierungsstellen. Methylierung verringert die Rezeptoraktivität (wirkt wie Ligandbindung) E.coli kann Aspartat von 10-10 M-10-3 M (über 8 Größenordnungen) in der Konzentration nachspüren. Bakterien reagieren auf eine schnelle zeitliche Änderung des Lockstoffes nicht auf die absolute Konzentration
Adaption unter Änderung der CheR Konzentration Die Adaptionspräzision ist unabhängig von CheR CheR wird mittels Plasmid unter externer Kontrolle eines Lac Operons von außen erhöht.
Adaption unter Änderung der CheR Konzentration Die Adaptionspräzision ist unabhängig von CheR CheR wird mittels Plasmid unter externer Kontrolle eines Lac Operons von außen erhöht.
Vergleich mit Systemkontrolle:Integral feedback control Doyl (1999) Der abgebildete Regelkreis verstärkt u in ein Ausgangssignal y. Dieses wird über den Integral feedback langsam auf Null zurückgeregelt (siehe Integralanteil bei PID Controllern). u: Eingangssignal (Lockstoffkonz.) x: Integral feedback (Grad der Methylierung) y0: Sollgröße y1-y0:Abweichung y: Ausgangsgröße (Rezeptoraktivität)
Das intrazelluläre Signal-Netzwerk Aus genetischen Untersuchungen sind vier cytoplasmatische Proteine CheA, CheW, CheY und CheZ bekannt, die die chemotaktischen Rezeptoren mit dem Geißelmotor verbinden. CheW: Hilfsfaktor CheA : Proteinkinase CheY : Response Regulator CheZ : Phosphatase-activating Protein Reguliert durch den Rezeptor transferiert CheA eine Phosphorylierung auf das kleine CheY Protein. CheY-p diffundiert zu den Motorproteinen der Flagellen und induziert den Übergang des Motors in die Taumelbewegung. (CheY-p = Ein/Ausschalter)
Die Signalübertragunsfunktion ist sehr steil Hill koeffizient 10
Die Signalübertragunsfunktion ist sehr steil Hill Equation Binding a ligand to a receptor is often enhanced if there are already other ligands present (cooperative binding). The Hill coefficient, provides a way to quantify this effect, initially used for O2-binding to Hemoglobin. θ - fraction of ligand binding sites filled [L] - ligand concentration Kd - dissociation constant Hill coefficient n. Describes the cooperativity of ligand binding: n > 1 - Positively cooperative reaction: Once one ligand molecule is bound to the enzyme, its affinity for other ligand molecules increases. n < 1 - Negatively cooperative reaction: Once one ligand molecule is bound to the enzyme, its affinity for other ligand molecules decreases. n = 1 - Noncooperative reaction: Independent binding to the receptor. Same as Langmuir equation. Hill koeffizient 10
Messung der Motor-Signalübertragung Cluzel et al.
Grün-fluoreszierendes Protein Green fluorescent protein (GFP) From jellyfish Aequorea victoria Die drei Aminosäuren bilden den GFP chromophore GFP kann gentechnisch mit anderen Proteinen fusioniert werden. Cluzel et al. haben CheY-P–GFP unter der Kontrolle eines Lac Promoters konstruiert.
Fluoreszenz-korrelation erlaubt die Eichung der Proteinkonzentration in der Zelle
Die Signalübertragunsfunktion ist steil Ein-Aus Schalter-Charakteristik, Verstärkungsfaktor 10 Gesamtverstärkung 60 (bei Front-end Verstärkung 3-5) Nur 0.1% Änderung der Rezeptorbesetzung Hill koeffizient 10
-> Slides from Talk of Tom Duke, Cavendish Laboratory, UK
Signal processing by clusters of membrane receptors Tom Duke Cavendish Laboratory Ian Graham Dennis Bray Anatomy Department University of Cambridge
Protein network unstimulated stimulated CW CCW
Protein network unstimulated stimulated adapted
Exact adaptation • Barkai & Leibler ‘97 • If modification enzyme Che B binds only to active receptors, steady-state activity is • independent of ambient concentration • Robust, exact adaptation signal response concentration activity
Exact adaptation • Barkai & Leibler ‘97 • If modification enzyme Che B binds only to active receptors, steady-state activity is • independent of ambient concentration • Robust, exact adaptation • Separation of time scales: • conformational transition 10-5 s ? • ligand release 10-3 s • phosphorylation 10-1 s response time • methylation 10 s adaptation time signal response concentration activity
Protein network unstimulated stimulated adapted Step 1: Introduction of two conformation states of the receptor: active and inactive Step 2: Let these conformations couple to neighboring receptors
Two-state ligand-binding protein • ligand binds more strongly to inactive state inactive (-) active (+) EA unbound -EA bound
Two-state ligand-binding protein • ligand binds more strongly to inactive state • mean activity depends on ligand concentration inactive (-) active (+) EA unbound -EA bound - Chemical potential of bound ligand Probability of inactive state Parallel to binding, receptors become inactive (-): Fast way to some sort of 'adaption' behaviour
Simple model:thermodynamics • for the symmetric case • the adapted system with P(si)=0.5 has Consider different receptors i with three states: -1 inactive +1 active si= unbound 0 unliganded 1 liganded li= 0 unmodified 1 modified mi= bound unmodified modified
Simple model:sensitivity to fractional change in concentration EL • Sensitivity of adaptedsystem change in ligand occupancy • depends on shape of ligand-binding curve Occupancy PL c/Kd
Model:sensitivity to fractional change in concentration • Sensitivity of adaptedsystem change in ligand occupancy • depends on shape of ligand-binding curve EL Sensitivity Occupancy PL c/Kd