Download
1 / 1
10 likes | 449 Views
0.3 nA 0.02 nA 1 nA 0.05 nA 0.5 nA 1 nA 0.05 nA 1 nA 0.5 nA 50 ms 50 ms 50 ms 50 ms 50 ms 50 ms 50 ms 50 ms 50 ms

E N D
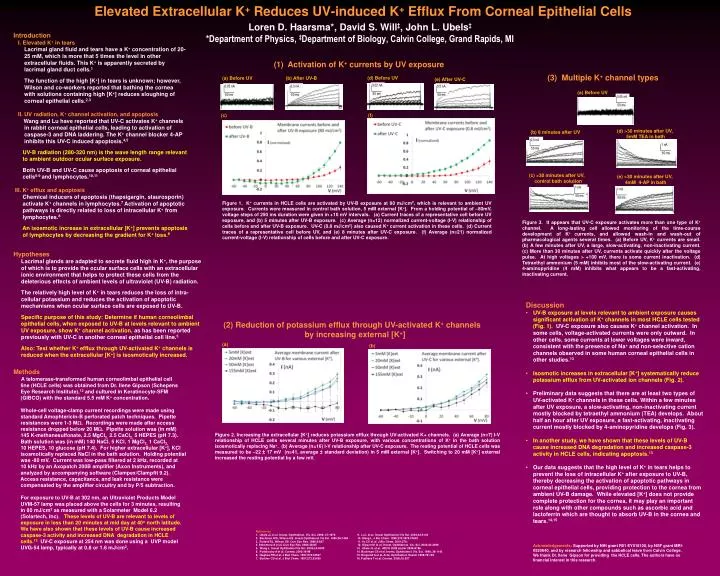
0.3 nA 0.02 nA 1 nA 0.05 nA 0.5 nA 1 nA 0.05 nA 1 nA 0.5 nA 50 ms 50 ms 50 ms 50 ms 50 ms 50 ms 50 ms 50 ms 50 ms Elevated Extracellular K+ Reduces UV-induced K+ Efflux From Corneal Epithelial CellsLoren D. Haarsma*, David S. Will‡, John L. Ubels‡ *Department of Physics, ‡Department of Biology, Calvin College, Grand Rapids, MI Introduction I. Elevated K+ in tears Lacrimal gland fluid and tears have a K+ concentration of 20- 25 mM, which is more that 5 times the level in other extracellular fluids. This K+ is apparently secreted by lacrimal gland duct cells.1 The function of the high [K+] in tears is unknown; however, Wilson and co-workers reported that bathing the cornea with solutions containing high [K+] reduces sloughing of corneal epithelial cells.2,3 II. UV radiation, K+ channel activation, and apoptosis Wang and Lu have reported that UV-C activates K+ channels in rabbit corneal epithelial cells, leading to activation of caspase-3 and DNA laddering. The K+ channel blocker 4-AP inhibits this UV-C induced apoptosis.4,5 UV-B radiation (280-320 nm) is the wave length range relevant to ambient outdoor ocular surface exposure. Both UV-B and UV-C cause apoptosis of corneal epithelial cells6-9 and lymphocytes.10,11 III. K+ efflux and apoptosis Chemical inducers of apoptosis (thapsigargin, staurosporin) activate K+ channels in lymphocytes.7 Activation of apoptotic pathways is directly related to loss of intracellular K+ from lymphocytes.8 An isosmotic increase in extracellular [K+] prevents apoptosis of lymphocytes by decreasing the gradient for K+ loss.8 (1) Activation of K+ currents by UV exposure (3) Multiple K+ channel types (d) Before UV (a) Before UV (b) After UV-B (e) After UV-C (a) Before UV (c) (f) (d) >30 minutes after UV, 5mM TEA in bath (b) 8 minutes after UV (c) >30 minutes after UV, control bath solution (e) >30 minutes after UV, 4mM 4-AP in bath Figure 3. It appears that UV-C exposure activates more than one type of K+ channel. A long-lasting cell allowed monitoring of the time-course development of K+ currents, and allowed wash-in and wash-out of pharmacological agents several times. (a) Before UV, K+ currents are small. (b) A few minutes after UV, a large, slow-activating, non-inactivating current. (c) More than 30 minutes after UV, currents activate quickly after the voltage pulse. At high voltages > +100 mV, there is some current inactivation. (d) Tetraethyl ammonium (5 mM) inhibits most of the slow-activating current. (e) 4-aminopyridine (4 mM) inhibits what appears to be a fast-activating, inactivating current. Figure 1. K+ currents in HCLE cells are activated by UV-B exposure at 80 mJ/cm2, which is relevant to ambient UV exposure. Currents were measured in control bath solution, 5 mM external [K+]. From a holding potential of –80mV, voltage steps of 250 ms duration were given in +10 mV intervals. (a) Current traces of a representative cell before UV exposure, and (b) 5 minutes after UV-B exposure. (c) Average (n=12) normalized current-voltage (I-V) relationship of cells before and after UV-B exposure. UV-C (0.8 mJ/cm2) also caused K+ current activation in these cells. (d) Current traces of a representative cell before UV, and (e) 8 minutes after UV-C exposure. (f) Average (n=21) normalized current-voltage (I-V) relationship of cells before and after UV-C exposure. Hypotheses Lacrimal glands are adapted to secrete fluid high in K+, the purpose of which is to provide the ocular surface cells with an extracellular ionic environment that helps to protect these cells from the deleterious effects of ambient levels of ultraviolet (UV-B) radiation. The relatively high level of K+ in tears reduces the loss of intra- cellular potassium and reduces the activation of apoptotic mechanisms when ocular surface cells are exposed to UV-B. Specific purpose of this study: Determine if human corneolimbal epithelial cells, when exposed to UV-B at levels relevant to ambient UV exposure, show K+ channel activation, as has been reported previously with UV-C in another corneal epithelial cell line.5 Also: Test whether K+ efflux through UV-activated K+ channels is reduced when the extracellular [K+] is isosmotically increased. Methods A telomerase-transformed human corneolimbal epithelial cell line (HCLE cells) was obtained from Dr. Ilene Gipson (Schepens Eye Research Institute),12 and cultured in Keratinocyte-SFM (GIBCO) with the standard 5.5 mM K+ concentration. Whole-cell voltage-clamp current recordings were made using standard Amophtericin-B perforated patch techniques. Pipette resistances were 1-3 MW. Recordings were made after access resistance dropped below 20 MW. Pipette solution was (in mM) 145 K-methanesulfonate, 2.5 MgCl2, 2.5 CaCl2, 5 HEPES (pH 7.3). Bath solution was (in mM) 140 NaCl, 5 KCl, 1 MgCl2, 1 CaCl2, 10 HEPES, 10 glucose (pH 7.4). For higher extracellular [K+], KCl isosmotically replaced NaCl in the bath solution. Holding potential was -80 mV. Current was low-pass filtered at 2 kHz, recorded at 10 kHz by an Axopatch 200B amplifier (Axon Instruments), and analyzed by accompanying software (Clampex/Clampfit 9.2). Access resistance, capacitance, and leak resistance were compensated by the amplifier circuitry and by P/5 subtraction. For exposure to UV-B at 302 nm, an Ultraviolet Products Model UVM-57 lamp was placed above the cells for 3 minutes, resulting in 80 mJ/cm2 as measured with a Solarmeter Model 6.2 (Solartech, Inc). These levels of UV-B are relevant to levels of exposure in less than 20 minutes at mid day at 40o north latitude. We have also shown that these levels of UV-B cause increased caspase-3 activity and increased DNA degradation in HCLE cells.13UV-C exposure at 254 nm was done using a UVP model UVG-54 lamp, typically at 0.8 or 1.6 mJ/cm2. • Discussion • UV-B exposure at levels relevant to ambient exposure causes significant activation of K+ channels in most HCLE cells tested (Fig. 1). UV-C exposure also causes K+ channel activation. In some cells, voltage-activated currents were only outward. In other cells, some currents at lower voltages were inward, consistent with the presence of Na+ and non-selective cation channels observed in some human corneal epithelial cells in other studies.13 • Isosmotic increases in extracellular [K+] systematically reduce potassium efflux from UV-activated ion channels (Fig. 2). • Preliminary data suggests that there are at least two types of UV-activated K+ channels in these cells. Within a few minutes after UV exposure, a slow-activating, non-inactivating current mostly blocked by tetraethyl ammonium (TEA) develops. About half an hour after UV exposure, a fast-activating, inactivating current mostly blocked by 4-aminopyridine develops (Fig. 3). • In another study, we have shown that these levels of UV-B cause increased DNA degradation and increased caspase-3 activity in HCLE cells, indicating apoptosis.13 • Our data suggests that the high level of K+ in tears helps to prevent the loss of intracellular K+ after exposure to UV-B, thereby decreasing the activation of apoptotic pathways in corneal epithelial cells, providing protection to the cornea from ambient UV-B damage. While elevated [K+] does not provide complete protection for the cornea, it may play an important role along with other compounds such as ascorbic acid and lactoferrin which are thought to absorb UV-B in the cornea and tears.14,15 (a) (b) (2) Reduction of potassium efflux through UV-activated K+ channels by increasing external [K+] Figure 2. Increasing the extracellular [K+] reduces potassium efflux through UV-activated K+ channels. (a) Average (n=7) I-V relationship of HCLE cells several minutes after UV-B exposure, with various concentrations of K+ in the bath solution isosmotically replacing Na+. (b) Average (n=16) I-V relationship after UV-C exposure. The resting potential of HCLE cells was measured to be –22 ± 17 mV (n=41, average ± standard deviation) in 5 mM external [K+]. Switching to 20 mM [K+] external increased the resting potential by a few mV. References 1. Ubels JL et al. Invest. Ophthalmol. Vis. Sci. 2006; 47:1876 9. Lu L et al. Invest Ophthalmol Vis Sci. 2003;44:5102 2. Bachman WG, Wilson GS. Invest Ophthalmol Vis Sci. 1985;26:1484 10. Wang L. J Bio Chem. 1999;274:3678 37602 3. Fullard RJ, Wilson GS. Curr Eye Res. 1986;5:847 11. Vu CC et al. J Bio Chem. 2001;276: 4. Shimmura S et al. Curr Eye Res. 2004;28:85 12. Gipson IK et al. Invest. Ophthalmol. Vis. Sci. 2003;44:2496 5. Wang L. Invest Ophthalmol Vis Sci. 2003;44:5095 13. Ubels JL et al. ARVO 2008 poster 2934/A186. 6. Podskochy A et al. Cornea. 2000;19:99 14. Bockman CS et al. Invest. Ophthalmol. Vis. Sci. 1998; 39:1143 6. Hughes FM et al. J Biol Chem. 1997;272:30567 15. Ringvold A et al. Acta. Ophthalmol. Scand. 1998;76:149 7. Bortner CD et al. J Biol Chem. 1997;272:32436 16. Fujihara T et al. Cornea. 2000;19:207 Acknowledgements: Supported by NIH grant R01 EY018100, by NSF grant MRI-0520840, and by research fellowship and sabbatical leave from Calvin College. We thank Dr. Ilene Gipson for providing the HCLE cells. The authors have no financial interest in this research.