Download
1 / 1

E N D
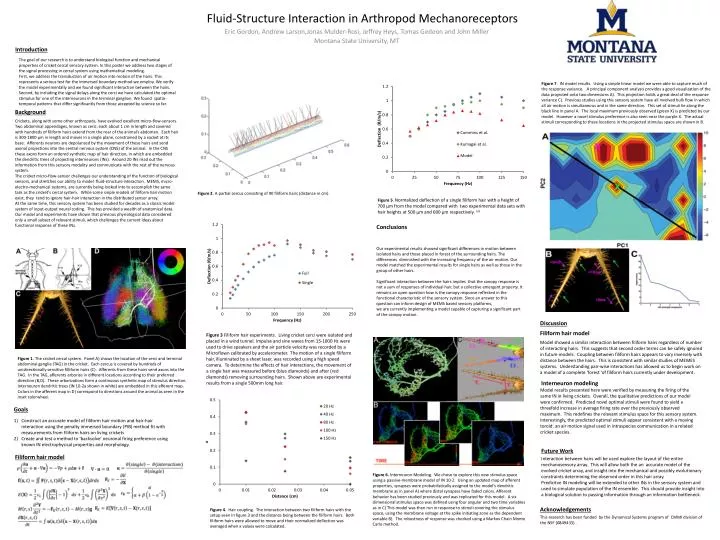
Figure 7.IN model results. Using a simple linear model we were able to capture much of the response variance. A principal component analysis provides a good visualization of the data projected onto two dimensions A). This projection holds a great deal of the response variance C). Previous studies using this sensory system have all involved bulk flow in which all air motion is simultaneous and in the same direction. This set of stimuli lie along the black line in panel A. The local maximum previously observed (green X) is predicted by our model. However a novel stimulus preference is also seen near the purple X. The actual stimuli corresponding to these locations in the projected stimulus space are shown in B. Introduction The goal of our research is to understand biological function and mechanical properties of cricket cercal sensory system. In this poster we address two stages of the signal processing in cercal system using mathematical modeling. First, we address the transduction of air motion into motion of the hairs. This represents a serious test for the immersed boundary method we employ. We verify the model experimentally and we found significant interaction between the hairs. Second, by including the signal delays along the cerci we have calculated the optimal stimulus for one of the interneurons in the terminal ganglion. We found spatio-temporal patterns that differ significantly from those accepted by science so far. Background Mechanosensor model Crickets, along with some other arthropods, have evolved excellent micro-flow-sensors. Two abdominal appendages, known as cerci, each about 1 cm in length and covered with hundreds of filiform hairs extend from the rear of the animal’s abdomen. Each hair is 300-1800 µm in length and moves in a single plane, constrained by a socket at its base. Afferents neurons are depolarized by the movement of these hairs and send axonal projections into the central nervous system (CNS) of the animal. In the CNS these axons form an ordered synthetic map of hair direction, in which are embedded the dendritic trees of projecting interneurons (INs). Around 20 INs read out the information from this sensory modality and communicate with the rest of the nervous system. The cricket micro-flow-sensor challenges our understanding of the function of biological sensors, and stretches our ability to model fluid-structure interaction. MEMS, micro-electro-mechanical systems, are currently being looked into to accomplish the same task as the cricket’s cercal system. While some sinple models of filiform hair motion exist, they tend to ignore hair-hair interaction in the distributed sensor array. At the same time, this sensory system has been studied for decades as a classic model system of input-output neural coding. This has provided a wealth of anatomical data. Our model and experiments have shown that previous physiological data considered only a small subset of relevant stimuli, which challenges the current ideas about functional response of these INs. Fluid-Structure Interaction in Arthropod Mechanoreceptors Figure 2. A partial cercus consisting of 90 filiform hairs (distance in cm). Interneuron modeling Model results presented here were verified by measuring the firing of the same IN in living crickets. Overall, the qualitative predictions of our model were confirmed. Predicted novel optimal stimuli were found to yield a threefold increase in average firing rate over the previously observed maximum. This redefines the relevant stimulus space for this sensory system. Interestingly, the predicted optimal stimuli appear consistent with a moving toroid: an air motion signal used in intraspecies communication in a related cricket species. Conclusions Figure 6. Interneuron Modeling. We chose to explore this new stimulus space using a passive-membrane model of IN 10-2. Using an updated map of afferent properties, synapses were probabilistically assigned to the model’s dendritic membrane as in panel A) where distal synapses have faded colors. Afferent behavior has been studied previously and was replicated for this model. A six dimensional stimulus space was defined using four angular and two time variables as in C) This model was then run in response to stimuli covering the stimulus space, using the membrane voltage at the spike initiating zone as the dependent variable B). The robustness of response was checked using a Markov Chain Monte Carlo method. Our experimental results showed significant differences in motion between isolated hairs and those placed in forest of the surrounding hairs. The differences diminished with the increasing frequency of the air motion. Our model matched the experimental results for single hairs as well as those in the group of other hairs. Significant interaction between the hairs implies that the canopy response is not a sum of responses of individual hair, but a collective emergent property. It remains an open question how is the canopy response reflected in the functional characteristic of the sensory system. Since an answer to this question can inform design of MEMS based sensory platforms, we are currently implementing a model capable of capturing a significant part of the canopy motion. Filiform hair model Model showed a similar interaction between filiform hairs regardless of number of interacting hairs. This suggests that second order terms can be safely ignored in future models. Coupling between filiform hairs appears to vary inversely with distance between the hairs. This is consistent with similar studies of MEMES systems. Understanding pair-wise interactions has allowed us to begin work on a model of a complete ‘forest ‘of filiform hairs currently under development. Eric Gordon, Andrew Larson,JonasMulder-Rosi, Jeffrey Heys, Tomas Gedeon and John Miller Montana State University, MT Discussion Figure 3 Filiform hair experiments. Living cricket cerci were isolated and placed in a wind tunnel. Impulse and sine waves from 15-1000 Hz were used to drive speakers and the air particle velocity was recorded by a Microflown calibrated by accelerometer. The motion of a single filiform hair, illuminated by a sheet laser, was recorded using a high speed camera. To determine the effects of hair interactions, the movement of a single hair was measured before (blue diamonds) and after (red diamonds) removing surrounding hairs. Shown above are experimental results from a single 500mm long hair. Future Work Interaction between hairs will be used explore the layout of the entire mechanosensory array. This will allow both the an accurate model of the evolved cricket array, and insight into the mechanical and possibly evolutionary constraints determining the observed order in this hair array. Predicitve IN modeling will be extended to other INs in the sensory system and used to simulate population of the IN ensemble. This should provide insight into a biological solution to passing information through an information bottleneck. Goals • Construct an accurate model of filiform hair motion and hair-hair interaction using the penalty immersed boundary (PIB) method fit with measurements from filiform hairs on living crickets • Create and test a method to ‘backsolve’ neuronal firing preference using known IN electrophysical properties and morphology. Filiform hair model Figure 1. The cricket cercal system. Panel A) shows the location of the cerci and terminal abdominal ganglia (TAG) in the cricket. Each cercus is covered by hundreds of unidirectionally sensitive filliform hairs (C). Afferents from these hairs send axons into the TAG. In the TAG, afferents arborize in different locations according to their preferred direction (B,D). These arborizations form a continuous synthetic map of stimulus direction. Interneuron dendritic trees (IN 10-2a shown in white) are embedded in this afferent map. Colors in the afferent map in D) correspond to directions around the animal as seen in the inset colorwheel. Acknowledgements Figure 5. Normalized deflection of a single filiform hair with a height of 700 µm from the model compared with two experimental data sets with hair heights at 500 µm and 600 µm respectively. 3,8 This research has been funded by the Dynamical Systems program of CMMI division of the NSF (0849433). Figure 4. Hair coupling. The interaction between two filiform hairs with the setup seen in figure 3 and the distance being between the filiform hairs. Both filiform hairs were allowed to move and their normalized deflection was averaged when κ values were calculated.